Særtrekk ved hvaler
11.05.2014 09:37Hvalens dypdykk kan vare en time:Saertrekk ved hvaler.pdf (591470)
Les merHvalens dypdykk kan vare en time:Saertrekk ved hvaler.pdf (591470)
Les merFugler er finjusterte dyr:Spesielle trekk ved fugler.pdf (523889)
Les merGrunntypemodell og menneskets stamtre: Homo erectus samme grunntype.pdf (428939)
Les merI Espen Ottosens bok: 'Hvorfor skriver ikke Gud på himmelen?'
bringer han videre noen innvendinger mot ikke-reduserbar kompleksitet.
Ved nærmere øyesyn, viser det seg at disse bygger mer på antagelser, enn realiteter:
Ikke-reduserbar-kompleksitet.pdf (440690)
Les merEn bevegelse i Isaac Newtons ånd
fra kap. 6 i 'Darwin 200 år-en festbrems; Peder A. Tyvand; Origo forlag. (Boka ikke til salgs lenger)
 Vi assosierer oss med bevegelsen Intelligent Design (ID). Denne bevegelsen er omgitt av avvisende uvenner og bedrevitende venner. Uvennene vil bekjempe oss, mens vennene vil belære oss. Uvennene og vennene er til sammen så dominerende at det ofte ikke blir plass til oss. Valget mellom å bli beskutt av uvenner og belært av venner er heller ikke enkelt.. Verken venner eller uvenner synes ha fått med seg at vi er en bevegelse som prøver blåse liv i våre lærefedres holdninger. Våre lærefedre er pionerene som sto bak den moderne naturvitenskapen, og blant dem var Isac Newton den fremste. Newton mente at naturen var skapt. Han mente naturlovene ga uttrykk for Skaperens orden og styring av naturen. Vi skal se nærmere på Newtons beskrivelse for den døde natur.
Vi assosierer oss med bevegelsen Intelligent Design (ID). Denne bevegelsen er omgitt av avvisende uvenner og bedrevitende venner. Uvennene vil bekjempe oss, mens vennene vil belære oss. Uvennene og vennene er til sammen så dominerende at det ofte ikke blir plass til oss. Valget mellom å bli beskutt av uvenner og belært av venner er heller ikke enkelt.. Verken venner eller uvenner synes ha fått med seg at vi er en bevegelse som prøver blåse liv i våre lærefedres holdninger. Våre lærefedre er pionerene som sto bak den moderne naturvitenskapen, og blant dem var Isac Newton den fremste. Newton mente at naturen var skapt. Han mente naturlovene ga uttrykk for Skaperens orden og styring av naturen. Vi skal se nærmere på Newtons beskrivelse for den døde natur. Darwinismen er ingen vitalisme. Dens grunnleggende ambisjon har hele tiden vært å forklare livet utelukkende ut fra prinsippene som gjelder i den døde naturen. De ønsker ikke å etablere ulike naturlover for levende og død natur, og har i minst et halvt århundre satset på at det ikke er noe skarpt skille mellom liv og død natur. Litt forenklet og lettere karikert har darwinismens syn på livet vært som følger: i) Liv er kompliserte og repeterende naturprosesser som involverer karbonatomer (C). ii) Grunnstoffet C er kun spesielt ved at det har valens 4, som setter det i stand til inntil 4 forbindelser med andre atomer. iii) Det som trengs for å danne liv er at C settes i forbindelse med tre andre grunnstoffer (H, O og N) i passende blanding. Under gunstige omstendigheter vil disse forbindelsene oppnå stor kompleksitet. 'Når kompleksiteten overstiger en viss grense, oppstår replikasjon (formering) som en naturlig ting. Vips, så er livet i gang. Det baller på seg videre, for egen maskin. Ved hjelp av mutasjoner og naturlig utvalg'.
Darwinismen er ingen vitalisme. Dens grunnleggende ambisjon har hele tiden vært å forklare livet utelukkende ut fra prinsippene som gjelder i den døde naturen. De ønsker ikke å etablere ulike naturlover for levende og død natur, og har i minst et halvt århundre satset på at det ikke er noe skarpt skille mellom liv og død natur. Litt forenklet og lettere karikert har darwinismens syn på livet vært som følger: i) Liv er kompliserte og repeterende naturprosesser som involverer karbonatomer (C). ii) Grunnstoffet C er kun spesielt ved at det har valens 4, som setter det i stand til inntil 4 forbindelser med andre atomer. iii) Det som trengs for å danne liv er at C settes i forbindelse med tre andre grunnstoffer (H, O og N) i passende blanding. Under gunstige omstendigheter vil disse forbindelsene oppnå stor kompleksitet. 'Når kompleksiteten overstiger en viss grense, oppstår replikasjon (formering) som en naturlig ting. Vips, så er livet i gang. Det baller på seg videre, for egen maskin. Ved hjelp av mutasjoner og naturlig utvalg'. a) Lovene for den døde natur er ikke-algoritmiske b) Lovene for livet er algoritmiske. Dette er ingen revitalisert vitalisme, der en postulerer et slags eksklusivt livsstoff i levende organismer. Skillet mellom det algoritmiske og det ikke-algoritmiske ikke-liv, går på mer abstrakt plan. Det går på det kvalitative, i form av naturlovene og deres egenart. En grunnleggende innsikt står fast:
a) Lovene for den døde natur er ikke-algoritmiske b) Lovene for livet er algoritmiske. Dette er ingen revitalisert vitalisme, der en postulerer et slags eksklusivt livsstoff i levende organismer. Skillet mellom det algoritmiske og det ikke-algoritmiske ikke-liv, går på mer abstrakt plan. Det går på det kvalitative, i form av naturlovene og deres egenart. En grunnleggende innsikt står fast: Her kommer en mulig villedning inn i bildet: Det er maktpåliggende for dagens darwinister at vi skal tro at naturlovene for den døde natur er algoritmiske. Men likevel bruker man oftest ikke ekte døde ting til å bekrefte dette. Når vi første gang lærer om varmelærens 2.lov i skolen, knuser vi kanskje et glass eller stikker hull på en ballong. Dette er eksempel på irreversible prosesser. Slike prosesser kan ikke gå baklengs i tid. Men tingene som er knust, er menneskeskapte ting. Dette er ting som er produsert algoritmisk. Derfor kan disse tingene virkelig gå i stykker. Prosesser som resulterer i irreversible prosesser, vil alltid gjelde døde ting med lånt algoritmisk design. Altså ting som i større eller mindre grad forutsetter algoritmer.
Her kommer en mulig villedning inn i bildet: Det er maktpåliggende for dagens darwinister at vi skal tro at naturlovene for den døde natur er algoritmiske. Men likevel bruker man oftest ikke ekte døde ting til å bekrefte dette. Når vi første gang lærer om varmelærens 2.lov i skolen, knuser vi kanskje et glass eller stikker hull på en ballong. Dette er eksempel på irreversible prosesser. Slike prosesser kan ikke gå baklengs i tid. Men tingene som er knust, er menneskeskapte ting. Dette er ting som er produsert algoritmisk. Derfor kan disse tingene virkelig gå i stykker. Prosesser som resulterer i irreversible prosesser, vil alltid gjelde døde ting med lånt algoritmisk design. Altså ting som i større eller mindre grad forutsetter algoritmer. 1) OM vi forsøker ødelegge en enkelt død ting, f.eks. en iskrystall, så kan en fullt likeverdig ting gjenskapes når betingelsene gjentar seg, f.eks. ved at en like fin iskrystall som den knuste fryses. Denne er blitt til ikke-algoritmisk, av eksakt det samme materialet. I dette laveste nivået foregår all bygging av strukturer ikke-algoritmisk. All ødeleggelse kan repareres ikke-algoritmisk. På dette nivået kan ting verken dø eller virkelig bli ødelagt.
1) OM vi forsøker ødelegge en enkelt død ting, f.eks. en iskrystall, så kan en fullt likeverdig ting gjenskapes når betingelsene gjentar seg, f.eks. ved at en like fin iskrystall som den knuste fryses. Denne er blitt til ikke-algoritmisk, av eksakt det samme materialet. I dette laveste nivået foregår all bygging av strukturer ikke-algoritmisk. All ødeleggelse kan repareres ikke-algoritmisk. På dette nivået kan ting verken dø eller virkelig bli ødelagt. 3. Levende vesener kan dø, f.eks. ei flue. Det vesenet som dør, lar seg normalt ikke gjenskape i samme form. En nytt individ av samme slag, kan bare bli til om artsfrender av den døde flua er i live. Det høyeste nivået er det eksklusive nivået for livet. Her foregår all bygging av strukturer ved hjelp av algoritmer. Ødeleggelse kan skje ikke-algoritmisk, ved at organismer dør og går i oppløsning. Men det nytter ikke bygge strukturer ikke-algoritmisk.
3. Levende vesener kan dø, f.eks. ei flue. Det vesenet som dør, lar seg normalt ikke gjenskape i samme form. En nytt individ av samme slag, kan bare bli til om artsfrender av den døde flua er i live. Det høyeste nivået er det eksklusive nivået for livet. Her foregår all bygging av strukturer ved hjelp av algoritmer. Ødeleggelse kan skje ikke-algoritmisk, ved at organismer dør og går i oppløsning. Men det nytter ikke bygge strukturer ikke-algoritmisk. Darwinismens forsøk på å forklare livets tilblivelse er ganske håpløst, fordi en starter på bar bakke med nivå 1 som eneste virkelighet. Om en utelukkende benytter seg av død materie og prosesser på nivå 1, vil nivå 2 befinne seg som en effektiv sperre mellom nivå 1 og nivå 3. Det er mulig darwinister vil prøve hoppe bukk over nivå 2, når de skal lage nivå 3 ut av nivå 1. Mer sannsynlig er det at de vil avvise hele problemstillingen. De føler jo selv at de sitter med svarene, men de er lite mottagelige for spørsmål. De fastholder det privilegiet det er å velge spørsmål som passer til svaret en sitter med. For svarene er alltid de samme: mutasjoner og naturlig utvalg.
Darwinismens forsøk på å forklare livets tilblivelse er ganske håpløst, fordi en starter på bar bakke med nivå 1 som eneste virkelighet. Om en utelukkende benytter seg av død materie og prosesser på nivå 1, vil nivå 2 befinne seg som en effektiv sperre mellom nivå 1 og nivå 3. Det er mulig darwinister vil prøve hoppe bukk over nivå 2, når de skal lage nivå 3 ut av nivå 1. Mer sannsynlig er det at de vil avvise hele problemstillingen. De føler jo selv at de sitter med svarene, men de er lite mottagelige for spørsmål. De fastholder det privilegiet det er å velge spørsmål som passer til svaret en sitter med. For svarene er alltid de samme: mutasjoner og naturlig utvalg.
 Livets algoritmer kommer fra liv. Dødens algoritmer eksisterer ikke. Darwinismens to hovedprinsipper er ikke algoritmiske i noen annen grad enn at de bryter ned algoritmer eller fjerner dem helt. Tilfeldige mutasjoner bryter ned livets algoritmer fra innsiden. Naturlig utvalg virker inn fra miljøet, og fjerner helt de algoritmene som ikke lykkes i konkurransen. Darwinismen i svakt modernisert utgave prøver å utgi både mutasjoner og naturlig utvalg for å være algoritmiske mekanismer. Disse tankene kan komme i to varianter, men begge er overflatiske:
Livets algoritmer kommer fra liv. Dødens algoritmer eksisterer ikke. Darwinismens to hovedprinsipper er ikke algoritmiske i noen annen grad enn at de bryter ned algoritmer eller fjerner dem helt. Tilfeldige mutasjoner bryter ned livets algoritmer fra innsiden. Naturlig utvalg virker inn fra miljøet, og fjerner helt de algoritmene som ikke lykkes i konkurransen. Darwinismen i svakt modernisert utgave prøver å utgi både mutasjoner og naturlig utvalg for å være algoritmiske mekanismer. Disse tankene kan komme i to varianter, men begge er overflatiske: Selv om det naturlig utvalg til tider er nådeløst, er det ikke algoritmisk. Til alt. 2 om at mutasjoner og naturlig utvalg til sammen skulle være algoritmiske, er flg. å si: Det ville i så fall si at mutasjoner utgjør formuleringsnivået for algoritmen, mens naturlig utvalg utgjør handlingsalternativet for den. Selv om det kan høres fint ut, blir det like meningsløst logisk som om en skulle spille sjakk med ludo-regler. Handlingsnivået for genetikken er individet og alle livsprosessene i individets kropp. Det naturlige utvalg handler utelukkende om omgivelsene og deres respons på individet. Seleksjon som prosess har ingenting å gjøre med iverksettelse av den genetiske algoritmen til et gitt individ.
Selv om det naturlig utvalg til tider er nådeløst, er det ikke algoritmisk. Til alt. 2 om at mutasjoner og naturlig utvalg til sammen skulle være algoritmiske, er flg. å si: Det ville i så fall si at mutasjoner utgjør formuleringsnivået for algoritmen, mens naturlig utvalg utgjør handlingsalternativet for den. Selv om det kan høres fint ut, blir det like meningsløst logisk som om en skulle spille sjakk med ludo-regler. Handlingsnivået for genetikken er individet og alle livsprosessene i individets kropp. Det naturlige utvalg handler utelukkende om omgivelsene og deres respons på individet. Seleksjon som prosess har ingenting å gjøre med iverksettelse av den genetiske algoritmen til et gitt individ. i tillegg til formulerings og handlingsnivået vi hittil har sett på. Det kan sammenlignes med hensikten i bondesjakk, å vinne spillet. For bondesjakk lar det seg greit gjøre å sette opp en algoritme til forsvar (Appendix 3 i boka). Men for det naturlige utvalg, henger det egoistiske hensiktsnivået i løse lufta. Vi kan riktignok observere dyrs egoistiske oppførsel, som ivaretar individets grunnleggende instinkter. På nåværende tidspunkt, aner vi ikke hvordan dette er lagt ned i individet. Uansett er det omgivelsene som bestemmer det naturlige utvalg. Genetikken hos et individ kan aldri bestemme sitt eget naturlige utvalg. Uansett hvilke intensjoner så dette individet måtte ha..
i tillegg til formulerings og handlingsnivået vi hittil har sett på. Det kan sammenlignes med hensikten i bondesjakk, å vinne spillet. For bondesjakk lar det seg greit gjøre å sette opp en algoritme til forsvar (Appendix 3 i boka). Men for det naturlige utvalg, henger det egoistiske hensiktsnivået i løse lufta. Vi kan riktignok observere dyrs egoistiske oppførsel, som ivaretar individets grunnleggende instinkter. På nåværende tidspunkt, aner vi ikke hvordan dette er lagt ned i individet. Uansett er det omgivelsene som bestemmer det naturlige utvalg. Genetikken hos et individ kan aldri bestemme sitt eget naturlige utvalg. Uansett hvilke intensjoner så dette individet måtte ha.. Intelligent Design (ID) avviker fra darwinismen ved at vi ikke aksepterer at destruktive krefter som tilfeldige mutasjoner og naturlig utvalg er tilstekkelig til å drive livet framover. Vi tror mer på en rekke konstruktive mekanismer og prinsipper:
Intelligent Design (ID) avviker fra darwinismen ved at vi ikke aksepterer at destruktive krefter som tilfeldige mutasjoner og naturlig utvalg er tilstekkelig til å drive livet framover. Vi tror mer på en rekke konstruktive mekanismer og prinsipper: Noe av det mest imponerende ved disse bakteriene, er at de benytter seg av perfekt rotasjon. Det er et sjeldent fenomen i naturen. Vi mennesker har ingen perfekte rotasjoner knyttet til vår kropp. Våre knær kan rotere omkring 150 grader, og halsen omtrent 180 grader. Noen fugler kan vri halsen trill rundt, altså 360 grader. Men det er bare én enkelt rotasjon. Med en perfekt rotasjon forstår vi her er rotasjon som kan gå rundt hvor mange ganger som helst. En rotasjon som gjentar seg selv uten noe reparasjonsproblem. Vanligvis tilegnes oppfinnelsen av hjulet perserne, ca 4000 år f.Kr. Men i dyreverdenen har det eksistert hjul så lenge nåværende type bakterier har eksistert (bildet).
Noe av det mest imponerende ved disse bakteriene, er at de benytter seg av perfekt rotasjon. Det er et sjeldent fenomen i naturen. Vi mennesker har ingen perfekte rotasjoner knyttet til vår kropp. Våre knær kan rotere omkring 150 grader, og halsen omtrent 180 grader. Noen fugler kan vri halsen trill rundt, altså 360 grader. Men det er bare én enkelt rotasjon. Med en perfekt rotasjon forstår vi her er rotasjon som kan gå rundt hvor mange ganger som helst. En rotasjon som gjentar seg selv uten noe reparasjonsproblem. Vanligvis tilegnes oppfinnelsen av hjulet perserne, ca 4000 år f.Kr. Men i dyreverdenen har det eksistert hjul så lenge nåværende type bakterier har eksistert (bildet). Rulling som bevegelsesform er ikke noe alternativ for dyr på landjorda. Når en struktur skal rulle rundt og rundt, blir det fort indre gnisninger som medfører behov for smøring. Det organ som best kan rulle hos mennesker, er øynene våre. De ruller i alle retninger, selv om vinkelen som øynene ruller kan være liten. Øynene smøres ved hjelp av tårevæske, og øyelokkene beskytter oss mot å få fremmedlegemer inn i tårvæsken. På denne bakgrunn er det oppsiktsvekkende at tarmbakterier er utstyrt med en liten framdriftsmotor, som praktiserer perfekt rulling. Motoren har en slags propell, som går rundt og rundt. Hvordan disse bakteriene løser 'smøreproblemet' er uvisst for oss. I likhet med oss mennesker, ser de ut til å ha utviklet noe i likhet med kulelager, for å minimalisere problemet.
Rulling som bevegelsesform er ikke noe alternativ for dyr på landjorda. Når en struktur skal rulle rundt og rundt, blir det fort indre gnisninger som medfører behov for smøring. Det organ som best kan rulle hos mennesker, er øynene våre. De ruller i alle retninger, selv om vinkelen som øynene ruller kan være liten. Øynene smøres ved hjelp av tårevæske, og øyelokkene beskytter oss mot å få fremmedlegemer inn i tårvæsken. På denne bakgrunn er det oppsiktsvekkende at tarmbakterier er utstyrt med en liten framdriftsmotor, som praktiserer perfekt rulling. Motoren har en slags propell, som går rundt og rundt. Hvordan disse bakteriene løser 'smøreproblemet' er uvisst for oss. I likhet med oss mennesker, ser de ut til å ha utviklet noe i likhet med kulelager, for å minimalisere problemet. Kroppsåpningenes intrikate design
Kroppsåpningenes intrikate design Ørene våre inneholder bevegelige deler. Små knokler som kalles for hammeren, stigbøylen og ambolten. Disse befinner seg i det indre øret, og er heldigvis beskyttet via trommehinnen. Øreknoklene er lenket sammen på en måte som gjør at de klarer seg med svært lite smøring. Virkemåten er fascinerende. Vi gjengir fra Utdanningsarkivet: Nervefibre inne i sneglehuset samler seg i hørselsnerven som bringer hørselssignaler til hjernen. Hørselssignaler oppstår ved at lyd setter trommehinnen i bevegelser. Disse bevegelsene overføres via ørebenskjeden (hammeren, ambolten og stigbøylen i mellomøret) til væsken i sneglehuset. Avhengig av frekvensen til lydbølgene, er det ulike nervefibre som registrerer lydimpulsen. De ulike nervefibrene transporterer altså ulike hørselsinntrykk som hjernen så kobler sammen til det lydbildet vi hører. Altså et intrikat samspill, som virker designet om en betrakter det med 'åpne øyne'?
Ørene våre inneholder bevegelige deler. Små knokler som kalles for hammeren, stigbøylen og ambolten. Disse befinner seg i det indre øret, og er heldigvis beskyttet via trommehinnen. Øreknoklene er lenket sammen på en måte som gjør at de klarer seg med svært lite smøring. Virkemåten er fascinerende. Vi gjengir fra Utdanningsarkivet: Nervefibre inne i sneglehuset samler seg i hørselsnerven som bringer hørselssignaler til hjernen. Hørselssignaler oppstår ved at lyd setter trommehinnen i bevegelser. Disse bevegelsene overføres via ørebenskjeden (hammeren, ambolten og stigbøylen i mellomøret) til væsken i sneglehuset. Avhengig av frekvensen til lydbølgene, er det ulike nervefibre som registrerer lydimpulsen. De ulike nervefibrene transporterer altså ulike hørselsinntrykk som hjernen så kobler sammen til det lydbildet vi hører. Altså et intrikat samspill, som virker designet om en betrakter det med 'åpne øyne'? Et særtrekk ved våre kroppsåpninger er at de har innerhud og ytterhud. Skillet mellom inner -og ytterhud er mer eller mindre ulikt skarpt for ulike åpninger. I munnen vår er dette skillet ganske kontinuerlig. Det går ingen klar grense mellom indre og ytre delen av leppene våre. De indre delene av leppene mottar beskyttelse i form av fuktighet hele tiden.. De ytterste delene er eksponert mot lufta, og vi har ingen automatisk refleks for å beskytte dem. Særlig om leppene er utsatt for mye tørr luft og sol, er vi nødt til å fukte leppene i blant for at de ikke skal sprekke opp. Det at våre kroppsåpninger trenger en form for smøring, har en sammenheng med interessante overganger mellom innerhud og ytterhud.
Et særtrekk ved våre kroppsåpninger er at de har innerhud og ytterhud. Skillet mellom inner -og ytterhud er mer eller mindre ulikt skarpt for ulike åpninger. I munnen vår er dette skillet ganske kontinuerlig. Det går ingen klar grense mellom indre og ytre delen av leppene våre. De indre delene av leppene mottar beskyttelse i form av fuktighet hele tiden.. De ytterste delene er eksponert mot lufta, og vi har ingen automatisk refleks for å beskytte dem. Særlig om leppene er utsatt for mye tørr luft og sol, er vi nødt til å fukte leppene i blant for at de ikke skal sprekke opp. Det at våre kroppsåpninger trenger en form for smøring, har en sammenheng med interessante overganger mellom innerhud og ytterhud. Ingen grense under vann
Ingen grense under vann i) Ingen grenser mellom fritt flytende proteiner og celler (med membran) ii) Ingen grenser mellom encellet og flercellet liv iii) ingen grense mellom de første dyr og de første planter iv) ingen grense mellom konsumenter og produsenter av organisk materiale (fotosyntesen) v) ingen grense mellom bløtdyr og virveldyr vi) ingen grense mellom livet i havet og det første liv på land vii) ingen grense mellom krypdyr og pattedyr viii) ingen grense mellom aper og mennesker
i) Ingen grenser mellom fritt flytende proteiner og celler (med membran) ii) Ingen grenser mellom encellet og flercellet liv iii) ingen grense mellom de første dyr og de første planter iv) ingen grense mellom konsumenter og produsenter av organisk materiale (fotosyntesen) v) ingen grense mellom bløtdyr og virveldyr vi) ingen grense mellom livet i havet og det første liv på land vii) ingen grense mellom krypdyr og pattedyr viii) ingen grense mellom aper og mennesker a) Krypdyr legger egg, pattedyr har livmor og føder levende unger. b) Krypdyr har eggeplomme, pattedyr har melkekjertler. c) Krypdyr har kloakkåpning, Pattedyr har separate kroppsåpninger for forplantning og avføring. d) Krypdyr har skjell, pattedyr har hud med hår. e) Krypdyr har fem kjeveledd på hver side, pattedyr har bare ett kjevebein på hver side. f) Krypdyr har primitiv hørsel, pattedyr har øreknokler som gir bedre hørselsbilde. g) Krypdyr er vekselvarme, pattedyr er varmblodige.
a) Krypdyr legger egg, pattedyr har livmor og føder levende unger. b) Krypdyr har eggeplomme, pattedyr har melkekjertler. c) Krypdyr har kloakkåpning, Pattedyr har separate kroppsåpninger for forplantning og avføring. d) Krypdyr har skjell, pattedyr har hud med hår. e) Krypdyr har fem kjeveledd på hver side, pattedyr har bare ett kjevebein på hver side. f) Krypdyr har primitiv hørsel, pattedyr har øreknokler som gir bedre hørselsbilde. g) Krypdyr er vekselvarme, pattedyr er varmblodige. Til sammenligning kan vi tenke på urfuglen (Archaeoteryx), og dens status som mellomform mellom krypdyr og fugler. Den var uten tvil en fugl: den hadde fjær og ikke skjell. Den hadde nebb og ingen kjeve. Den var ganske sikkert varmblodig og ikke vekselvarm. Den hadde vinger med halefjær, som den flakset med når den fløy. Intet krypdyr har hatt fjær eller fuglevinger. Flyveøgler hadde svevehud, i likhet med flaggermus. Det var to ting som fikk 'urfuglen' til å ligne på et krypdyr: Det ene var at den hadde masse små tenner. Det andre at den hadde klør på vingene. Til tross for sine små likheter med krypdyr, har Archaeoteryx en ikonisk status i darwinismen. Den betraktes som et 'endegyldig bevis' på overgangen mellom krypdyr og fugler. Den visker liksom ut alle grenser og gjør overgangen mellom krypdyr og fugler til en 'naturlig og hverdagslig' sak.
Til sammenligning kan vi tenke på urfuglen (Archaeoteryx), og dens status som mellomform mellom krypdyr og fugler. Den var uten tvil en fugl: den hadde fjær og ikke skjell. Den hadde nebb og ingen kjeve. Den var ganske sikkert varmblodig og ikke vekselvarm. Den hadde vinger med halefjær, som den flakset med når den fløy. Intet krypdyr har hatt fjær eller fuglevinger. Flyveøgler hadde svevehud, i likhet med flaggermus. Det var to ting som fikk 'urfuglen' til å ligne på et krypdyr: Det ene var at den hadde masse små tenner. Det andre at den hadde klør på vingene. Til tross for sine små likheter med krypdyr, har Archaeoteryx en ikonisk status i darwinismen. Den betraktes som et 'endegyldig bevis' på overgangen mellom krypdyr og fugler. Den visker liksom ut alle grenser og gjør overgangen mellom krypdyr og fugler til en 'naturlig og hverdagslig' sak. Darwinister prøver få seg selv og andre til å akseptere at alle skarpe grenser, er noe som naturen har funnet på senere. Vesener som er litt det ene og litt det andre, er 'ulne'. En ullen livsform uten presise spesifikasjoner, har ikke mer enn en liksom-overlevelse. Om dette gjelder de fleste livsformer i omgivelsene, overser man en helt avgjørende sak: 'Å være levedyktig er viktigere enn å hevde seg i konkurransen.' Fordi konkurransen visstnok var så svak 'en gang i fortiden', ser en helt bort fra alle fint avstemte algoritmiske mekanismer som må samvirke, om et individ i det hele tatt skal klare seg. Overlevelse synes være et lite problem for dem, i forhold til det å konkurrere med de andre livsformene. Men naturlig utvalg velger, som før nevnt, bare bort ting. Når en velger noe, velger en samtidig vekk noe annet. En som skal gifte seg, velger éi dame og samtidig bort de andre..
Darwinister prøver få seg selv og andre til å akseptere at alle skarpe grenser, er noe som naturen har funnet på senere. Vesener som er litt det ene og litt det andre, er 'ulne'. En ullen livsform uten presise spesifikasjoner, har ikke mer enn en liksom-overlevelse. Om dette gjelder de fleste livsformer i omgivelsene, overser man en helt avgjørende sak: 'Å være levedyktig er viktigere enn å hevde seg i konkurransen.' Fordi konkurransen visstnok var så svak 'en gang i fortiden', ser en helt bort fra alle fint avstemte algoritmiske mekanismer som må samvirke, om et individ i det hele tatt skal klare seg. Overlevelse synes være et lite problem for dem, i forhold til det å konkurrere med de andre livsformene. Men naturlig utvalg velger, som før nevnt, bare bort ting. Når en velger noe, velger en samtidig vekk noe annet. En som skal gifte seg, velger éi dame og samtidig bort de andre.. Alle biologer er enige om at det skjer tilfeldige mutasjoner, og at det store flertall av dem er skadelige. Det er også enighet om at det finnes noen få mutasjoner som ikke umiddelbart ser ut til å være skadelige. Uenigheten mellom darwinister og ikke-darwinister på dette punktet dreier seg om hvorvidt det finnes gunstige mutasjoner. Selv om darwinister bare ønsker å svare ja på dette spørsmålet og legge det bak seg, skal vi imidlertid rippe litt opp i dette når vi har anledning. Først: for darwinister går dette spørsmålet 'på livet løs'. Et annet svar enn et ubetinget 'ja', kan ikke tolereres av dem. Om en føyer dem i det, blir alt bare idyll for dem: 'de gunstige mutasjonene hoper seg opp, sakte og sikkert. Seleksjon gjør at et mangfold av rikere og stadig bedre utvalg etablerer seg.' Men denne forestillingen trenger utfordres: 'Hvor blir det av alle de skadelige mutasjonene?
Alle biologer er enige om at det skjer tilfeldige mutasjoner, og at det store flertall av dem er skadelige. Det er også enighet om at det finnes noen få mutasjoner som ikke umiddelbart ser ut til å være skadelige. Uenigheten mellom darwinister og ikke-darwinister på dette punktet dreier seg om hvorvidt det finnes gunstige mutasjoner. Selv om darwinister bare ønsker å svare ja på dette spørsmålet og legge det bak seg, skal vi imidlertid rippe litt opp i dette når vi har anledning. Først: for darwinister går dette spørsmålet 'på livet løs'. Et annet svar enn et ubetinget 'ja', kan ikke tolereres av dem. Om en føyer dem i det, blir alt bare idyll for dem: 'de gunstige mutasjonene hoper seg opp, sakte og sikkert. Seleksjon gjør at et mangfold av rikere og stadig bedre utvalg etablerer seg.' Men denne forestillingen trenger utfordres: 'Hvor blir det av alle de skadelige mutasjonene? De skadelige mutasjonene var jo i stort flertall. De utgjorde baksiden av medaljen: det var nødvendig å tillate mange skadelige mutasjoner for at det skulle dukke opp noen få som ikke var det. I darwinistisk forestillingsverden forsvinner disse greit, på grunn av det naturlige utvalg. Det er imidlertid en fordreining og forflatning av virkeligheten. Det er to grunner til det: i) Det er mye vanskeligere å bli kvitt skadelige mutasjoner enn å samle opp evt. gunstige mutasjoner. Det skyldes nettopp at det store flertall mutasjoner er skadelige. ii) Hovedproblemet er at skadelige mutasjoner er recessive, og dermed gjemmer seg bort i populasjonen på kort sikt. Det må gjøres et møysommelig 'avlsarbeid' for å luke bort skadelige mutasjoner som hoper seg opp.
De skadelige mutasjonene var jo i stort flertall. De utgjorde baksiden av medaljen: det var nødvendig å tillate mange skadelige mutasjoner for at det skulle dukke opp noen få som ikke var det. I darwinistisk forestillingsverden forsvinner disse greit, på grunn av det naturlige utvalg. Det er imidlertid en fordreining og forflatning av virkeligheten. Det er to grunner til det: i) Det er mye vanskeligere å bli kvitt skadelige mutasjoner enn å samle opp evt. gunstige mutasjoner. Det skyldes nettopp at det store flertall mutasjoner er skadelige. ii) Hovedproblemet er at skadelige mutasjoner er recessive, og dermed gjemmer seg bort i populasjonen på kort sikt. Det må gjøres et møysommelig 'avlsarbeid' for å luke bort skadelige mutasjoner som hoper seg opp. Grunnen til at dette er vanskelig er at seleksjon ikke virker på en skadelig mutasjon, dersom denne er recessiv. Recessiv innebærer at det genetiske innhold ikke kommer til uttrykk i organismen, fordi andre gener er dominerende -og undertrykker det genetiske innhold i mutasjonen. Hver skadelige recessiv mutasjon må fortrinnsvis lukes ut ved tilfeldig drift i små populasjoner. Da kan en fort risikere at nyttige egenskaper forsvinner ut i lag med skadelige. De recessive mutasjonene blir det dermed veldig vanskelig å bli kvitt.
Grunnen til at dette er vanskelig er at seleksjon ikke virker på en skadelig mutasjon, dersom denne er recessiv. Recessiv innebærer at det genetiske innhold ikke kommer til uttrykk i organismen, fordi andre gener er dominerende -og undertrykker det genetiske innhold i mutasjonen. Hver skadelige recessiv mutasjon må fortrinnsvis lukes ut ved tilfeldig drift i små populasjoner. Da kan en fort risikere at nyttige egenskaper forsvinner ut i lag med skadelige. De recessive mutasjonene blir det dermed veldig vanskelig å bli kvitt. Det finnes også dominante mutasjoner som er skadelige. Over tid går det greit å bli kvitt disse. I slike tilfeller pleier en si at det naturlige utvalg har en 'konserverende effekt'. Det innebærer at det naturlige utvalg beskytter populasjonen mot å bli infisert av en negativ mutasjon. Men det er aldri snakk om at naturlig utvalg bringer noe nytt inn i populasjonen. Den konserverende effekten er en bivirkning av at de skadelige dominante mutasjonene velges bort. For å komme særlig videre med spørsmålet vi baler med, er det nødvendig å reformulere og presisere det til mer moderne ordbruk: 'Finnes det algoritmisk gunstige mutasjoner, som kan bygge opp en rikere og mer robust struktur i arvestoffet?'
Det finnes også dominante mutasjoner som er skadelige. Over tid går det greit å bli kvitt disse. I slike tilfeller pleier en si at det naturlige utvalg har en 'konserverende effekt'. Det innebærer at det naturlige utvalg beskytter populasjonen mot å bli infisert av en negativ mutasjon. Men det er aldri snakk om at naturlig utvalg bringer noe nytt inn i populasjonen. Den konserverende effekten er en bivirkning av at de skadelige dominante mutasjonene velges bort. For å komme særlig videre med spørsmålet vi baler med, er det nødvendig å reformulere og presisere det til mer moderne ordbruk: 'Finnes det algoritmisk gunstige mutasjoner, som kan bygge opp en rikere og mer robust struktur i arvestoffet?' den hva som bor i den. Det kommer da for en dag at mutasjonen er dødelig. Dødeligheten gir et klart bevis på at sigdcelle-mutasjonen er en algoritmisk ugunstig mutasjon.
den hva som bor i den. Det kommer da for en dag at mutasjonen er dødelig. Dødeligheten gir et klart bevis på at sigdcelle-mutasjonen er en algoritmisk ugunstig mutasjon. Blodets evne til å størkne, koagulering, er veldig viktig for oss mennesker. Denne evnen reduserer blodtap, beskytter sår og hjelper dem til å gro. Noen av oss er blødere. Det skyldes en genetisk defekt som er arvelig, og antagelig skyldes mutasjoner. Hos blødere kan ikke blodet levre seg (størkne) som normal. Vårt immunforsvar har til oppgave å forsvare kroppen mot ytre og indre fiender. Indre fiender kan være tidlige stadier av kreft. Noen ganger overreagerer immunsystemet og forårsaker betennelser, lammelser etc. Det kan kanskje være bedre enn at immunapparatet ikke reagerer i det hele tatt.
Blodets evne til å størkne, koagulering, er veldig viktig for oss mennesker. Denne evnen reduserer blodtap, beskytter sår og hjelper dem til å gro. Noen av oss er blødere. Det skyldes en genetisk defekt som er arvelig, og antagelig skyldes mutasjoner. Hos blødere kan ikke blodet levre seg (størkne) som normal. Vårt immunforsvar har til oppgave å forsvare kroppen mot ytre og indre fiender. Indre fiender kan være tidlige stadier av kreft. Noen ganger overreagerer immunsystemet og forårsaker betennelser, lammelser etc. Det kan kanskje være bedre enn at immunapparatet ikke reagerer i det hele tatt.
 Selv hvor frustrerende dette kan være, så er det også lærerikt. Det er en snodig inkonsekvens: darwinister kan og vil bruke sin overlegne posisjon i folks omdømme, enten de har gode argumenter eller ikke. Om ikke annet, fokuserer de på at darwinister er i flertall og preger mediedebatten. Så mange kan ikke ta feil, eller hva? Som vi innledningsvis var inne på, er det ikke et lite antall som går mot darwinismen av ulike grunner heller. At det her mer er spørsmål om kvalitet enn kvantitet, er ikke noe motstanderne ønsker vi skal videreformidle. De ønsker å være vinnere av maktkampen og vinnere av ensidige debatter.
Selv hvor frustrerende dette kan være, så er det også lærerikt. Det er en snodig inkonsekvens: darwinister kan og vil bruke sin overlegne posisjon i folks omdømme, enten de har gode argumenter eller ikke. Om ikke annet, fokuserer de på at darwinister er i flertall og preger mediedebatten. Så mange kan ikke ta feil, eller hva? Som vi innledningsvis var inne på, er det ikke et lite antall som går mot darwinismen av ulike grunner heller. At det her mer er spørsmål om kvalitet enn kvantitet, er ikke noe motstanderne ønsker vi skal videreformidle. De ønsker å være vinnere av maktkampen og vinnere av ensidige debatter. -4: Null eksistens for ID/kreasjonisme. Darwinistene oppfatter seg taktisk eller debattmessig å være alene. Derfor benytter de sin posisjon til å late som de er det, ved å drive monoton enetale. Motstandere er liksom utradert, og eksisterer ikke. 'Debatten' på diskusjonsfora eller i media består i at darwinister snakker med seg selv eller andre darwinister. Dette er den mest ytterliggående form for realitetsbenektelse. Selv de snevreste marxistiske diktaturer erkjente at det fantes 'klassefiender'. Likevel er debattnivået reelt i norsk skapelsesdebatt. Noen av våre bedrevitende 'venner' har forskanset seg der. Fagbiologiske motstandere kan derimot avholde seg fra dette absolutte lavmålet.
-4: Null eksistens for ID/kreasjonisme. Darwinistene oppfatter seg taktisk eller debattmessig å være alene. Derfor benytter de sin posisjon til å late som de er det, ved å drive monoton enetale. Motstandere er liksom utradert, og eksisterer ikke. 'Debatten' på diskusjonsfora eller i media består i at darwinister snakker med seg selv eller andre darwinister. Dette er den mest ytterliggående form for realitetsbenektelse. Selv de snevreste marxistiske diktaturer erkjente at det fantes 'klassefiender'. Likevel er debattnivået reelt i norsk skapelsesdebatt. Noen av våre bedrevitende 'venner' har forskanset seg der. Fagbiologiske motstandere kan derimot avholde seg fra dette absolutte lavmålet. -3: Darwinister lager stråmenn. Darwinister erkjenner at motstandere finnes. De påtår seg oppgaven å forklare offentligheten hva ID-bevegelsen står for. Vi spørres eller tillates ikke å gjøre det selv. Det er akkurat som en redd for hva vi kan komme til å si. Media vil først plassere oss i en bås. Når de så har fått merket ut, gjerne en karikatur, av hva ID-bevegelsen står for, går en til angrep på denne karikaturen. En kjent Oslo-professor har i fortiden stadig benyttet denne usaklige formen for liksom-debatt. Han sier først hva vi mener, så hva han selv mener om det han mener at vi mener. Det er ikke overraskende at han ofte har vunnet debatter med seg selv som eneste deltager.
-3: Darwinister lager stråmenn. Darwinister erkjenner at motstandere finnes. De påtår seg oppgaven å forklare offentligheten hva ID-bevegelsen står for. Vi spørres eller tillates ikke å gjøre det selv. Det er akkurat som en redd for hva vi kan komme til å si. Media vil først plassere oss i en bås. Når de så har fått merket ut, gjerne en karikatur, av hva ID-bevegelsen står for, går en til angrep på denne karikaturen. En kjent Oslo-professor har i fortiden stadig benyttet denne usaklige formen for liksom-debatt. Han sier først hva vi mener, så hva han selv mener om det han mener at vi mener. Det er ikke overraskende at han ofte har vunnet debatter med seg selv som eneste deltager. -2: Darwinistisk blindebukk-lek: ID-tilhengere/kreasjonister får slippe til, men ser eller hører ingenting til sine meningsmotstandere, så lenge meningsytringer foregår. En kan bli intervjuet av en journalist, så blir opptak kryssklippet og ferdig-redigert. Deretter blir darwinisten invitert inn. De får sitte og slutt-kommentere det redigerte opptaket av det vi har sagt. Så blir dette servert til offentligheten, uten noen mulighet til korreksjon/spesifisering fra vår side. Visse biologiprofessorer og mange norske media foretrekker denne debattformen. Erik Tunstad har brukt dette nivået flere ganger når forfatter her har vært intervjuet på NRK P2.
-2: Darwinistisk blindebukk-lek: ID-tilhengere/kreasjonister får slippe til, men ser eller hører ingenting til sine meningsmotstandere, så lenge meningsytringer foregår. En kan bli intervjuet av en journalist, så blir opptak kryssklippet og ferdig-redigert. Deretter blir darwinisten invitert inn. De får sitte og slutt-kommentere det redigerte opptaket av det vi har sagt. Så blir dette servert til offentligheten, uten noen mulighet til korreksjon/spesifisering fra vår side. Visse biologiprofessorer og mange norske media foretrekker denne debattformen. Erik Tunstad har brukt dette nivået flere ganger når forfatter her har vært intervjuet på NRK P2. -1: Darwinistisk debattregi: Vi møter darwinister i debatter som skal være frie, men preges av darwinist-vennlige debattledere. Alle sportsinteresserte vet at det er vanskelig å vinne, om dommeren konsekvent er med det andre laget. Parallellen er at det ofte er mindre kjente lag som utsettes for dette. I fotball klager motstandere av RBK ofte over det i forbindelse med utelatte straffer etc. Såkalte vennlige norske media befinner seg på dette nivået. De fleste er ikke fullt så vennlige. Denne forfatteren har opplevd et par ganger at NRK har vært på dette nivået, på direktesendte debatter. Om det er opptak, flyttes debatten ned til -2 nivået.
-1: Darwinistisk debattregi: Vi møter darwinister i debatter som skal være frie, men preges av darwinist-vennlige debattledere. Alle sportsinteresserte vet at det er vanskelig å vinne, om dommeren konsekvent er med det andre laget. Parallellen er at det ofte er mindre kjente lag som utsettes for dette. I fotball klager motstandere av RBK ofte over det i forbindelse med utelatte straffer etc. Såkalte vennlige norske media befinner seg på dette nivået. De fleste er ikke fullt så vennlige. Denne forfatteren har opplevd et par ganger at NRK har vært på dette nivået, på direktesendte debatter. Om det er opptak, flyttes debatten ned til -2 nivået. 0: En saklig og nyansert debatt, med programledere/regissører som konsekvent prøver være objektive. I deler av debatten kan det være umulig i praksis, men poenget er at helheten skal være balansert og objektiv. Det er ingen lett sak, selv om det ikke er umulig. Etter episode 1 synes det som valg av deltakere i programserien 'Gud og vitenskapen', et engelskprodusert serie som sendes på NRK 2 i januar 2014, prøver holde seg til denne standarden. Det skal bli spennende å følge det videre når temaet blir evolusjonslæren. Riktignok har NRK plassert det såpass sent på kveldstid at folk i full jobb, særlig pendlere velger det bort av den grunn. Men det finnes opptaksmuligheter..
0: En saklig og nyansert debatt, med programledere/regissører som konsekvent prøver være objektive. I deler av debatten kan det være umulig i praksis, men poenget er at helheten skal være balansert og objektiv. Det er ingen lett sak, selv om det ikke er umulig. Etter episode 1 synes det som valg av deltakere i programserien 'Gud og vitenskapen', et engelskprodusert serie som sendes på NRK 2 i januar 2014, prøver holde seg til denne standarden. Det skal bli spennende å følge det videre når temaet blir evolusjonslæren. Riktignok har NRK plassert det såpass sent på kveldstid at folk i full jobb, særlig pendlere velger det bort av den grunn. Men det finnes opptaksmuligheter.. som uvitenskapelig fordi det står i Bibelen.' De fleste biologiske fakta tilsier en overnaturlig skapelse, men vi vet lite konkret hvordan det skjedde. De første to kap. i Bibelen skildrer den på to ulike måter, hvor både menneskets guddommelige og jordiske side kommer fram.
som uvitenskapelig fordi det står i Bibelen.' De fleste biologiske fakta tilsier en overnaturlig skapelse, men vi vet lite konkret hvordan det skjedde. De første to kap. i Bibelen skildrer den på to ulike måter, hvor både menneskets guddommelige og jordiske side kommer fram. En har erstattet et dogme om overnaturlig skapelse, med et annet om 'naturlig tilblivelse' av seg selv. Naturalismen som livssyn var tilstede lenge før darwinismen. Darwins bidrag var liksom å gjøre dette synet tilforlatelig. Når han med sin gode fantasi kom opp med forklaringer om mutasjoner og naturlig utvalg, så var grunnlaget lagt. Det som trengtes var en herold til å spre budskapet, her gjorde aviser en viktig jobb. Og ateister og andre som ikke syntes noe om følgene av Gudstroen jublet. Tegning av menneskets avstamning ble gjort lenge før Darwin, og han fulgte opp med en ideologi av 'spekulasjoner'. Bevis i form av fossiler etc, regnet han med kom senere..
En har erstattet et dogme om overnaturlig skapelse, med et annet om 'naturlig tilblivelse' av seg selv. Naturalismen som livssyn var tilstede lenge før darwinismen. Darwins bidrag var liksom å gjøre dette synet tilforlatelig. Når han med sin gode fantasi kom opp med forklaringer om mutasjoner og naturlig utvalg, så var grunnlaget lagt. Det som trengtes var en herold til å spre budskapet, her gjorde aviser en viktig jobb. Og ateister og andre som ikke syntes noe om følgene av Gudstroen jublet. Tegning av menneskets avstamning ble gjort lenge før Darwin, og han fulgte opp med en ideologi av 'spekulasjoner'. Bevis i form av fossiler etc, regnet han med kom senere.. Kristen darwinister er nok en broket gruppe, når det kommer til stykket. De omfatter alt fra de som tror på 'gudfeldigheter' (tilsynelatende tilfeldigheter), til de som 'kobler ut' trossiden og stringent holder seg til det de oppfatter som 'bevis'. Skal en være konsekvent her, må det sies at om noen ikke holder seg konsekvent etter darwinismens tilfeldigheter som virkelige, ikke bare tilsynelatende tilfeldigheter, så åpner en opp for en Intelligens. 'Ja takk', til to logisk motstridende ting, er ikke særlig god filosofi. At intelligens skal anerkjennes av andre vitenskaper, men ikke her i slike engangshendinger virker som sagt merkelig. Nok om det, vi får forholde oss til det de sier de er: kristen-darwinister.
Kristen darwinister er nok en broket gruppe, når det kommer til stykket. De omfatter alt fra de som tror på 'gudfeldigheter' (tilsynelatende tilfeldigheter), til de som 'kobler ut' trossiden og stringent holder seg til det de oppfatter som 'bevis'. Skal en være konsekvent her, må det sies at om noen ikke holder seg konsekvent etter darwinismens tilfeldigheter som virkelige, ikke bare tilsynelatende tilfeldigheter, så åpner en opp for en Intelligens. 'Ja takk', til to logisk motstridende ting, er ikke særlig god filosofi. At intelligens skal anerkjennes av andre vitenskaper, men ikke her i slike engangshendinger virker som sagt merkelig. Nok om det, vi får forholde oss til det de sier de er: kristen-darwinister. Når det er sådd tvil om alle virkelig er darwinister i ordets stringente betydning, må det også sies at de har et annet forhold til den dominerende darwinismen. De er jo liksom begge deler, og da får ateister også et mer positivt syn på dem. Det er lett å ta kreasjonister på uforklarlige ting i Bibel i forhold til naturen, men når kristendarwinister leser om skapelsen blir det billedlig, at Gud står bak den. 'Bibelen svarer på hvorfor Gud skapte, mens vitenskapen på hvordan han gjorde det.' Det kan være fristende å være kristen-darwinist for å slippe motstand, skaffe seg spalteplass og kunne bruke bibelsitater uten å beskyldes for å fremme teologi i stedet for vitenskap.
Når det er sådd tvil om alle virkelig er darwinister i ordets stringente betydning, må det også sies at de har et annet forhold til den dominerende darwinismen. De er jo liksom begge deler, og da får ateister også et mer positivt syn på dem. Det er lett å ta kreasjonister på uforklarlige ting i Bibel i forhold til naturen, men når kristendarwinister leser om skapelsen blir det billedlig, at Gud står bak den. 'Bibelen svarer på hvorfor Gud skapte, mens vitenskapen på hvordan han gjorde det.' Det kan være fristende å være kristen-darwinist for å slippe motstand, skaffe seg spalteplass og kunne bruke bibelsitater uten å beskyldes for å fremme teologi i stedet for vitenskap. At tilhengere av Intelligent Design (ID) ikke har sluppet tilnærmelsesvis så mye til, kan være fordi bevegelsen er relativt ny. Men en underliggende skepsis ligger der og dirrer: Egentlig er det teologi de er opptatt av, ikke vitenskap. Vi ber dermed kreasjonister om forståelse fordi vi ikke bruker bibelsitater så mye i denne debatten som kristen-darwinister. Det er nok også en historisk forklaring at historisk-kritisk metode fikk et større gjennomslag i Tyskland, der norsk luthersk kristendom har sin opprinnelse, enn i England der ca halvparten tror på evolusjonsteorien. I hvert fall gjelder det innenfor geisteligheten, og kristne undervisnings-institusjoner i Norge i dag.
At tilhengere av Intelligent Design (ID) ikke har sluppet tilnærmelsesvis så mye til, kan være fordi bevegelsen er relativt ny. Men en underliggende skepsis ligger der og dirrer: Egentlig er det teologi de er opptatt av, ikke vitenskap. Vi ber dermed kreasjonister om forståelse fordi vi ikke bruker bibelsitater så mye i denne debatten som kristen-darwinister. Det er nok også en historisk forklaring at historisk-kritisk metode fikk et større gjennomslag i Tyskland, der norsk luthersk kristendom har sin opprinnelse, enn i England der ca halvparten tror på evolusjonsteorien. I hvert fall gjelder det innenfor geisteligheten, og kristne undervisnings-institusjoner i Norge i dag. At kristen-darwinistene omfavnes på en annen måte enn tilhengere av Intelligent Design, gjør nok sitt til at kristen-darwinister gjerne også tror seg bedre i stand til å hevde og forsvare kristne interesser overfor ateister. Noen avisredaksjoner synes også være av den oppfatning at det er nok at kristen-darwinister slipper til. Noen av dem sukrer da den bitre pillen ved å si at det er til gagn for kristenheten at de fører debatten for oss. Og det til tross for at de står omtrent for det motsatte av oss i denne saken. Det eneste positive jeg er i stand til å se, er at motsetninger kan skape en spenning og en debatt som gjør spørsmål om Guds rolle i verden mer sentral. Det vil jo ikke være en liten betydning, og det ville faktisk være verdt prisen å måtte stille i annen rekke ved diskusjoner etc.Kristen-darwinister hevder gjerne at vi står for en Bibelforståelse, som ikke griper Bibelens intensjoner. Det er jo lett å se for seg at Bibelen ikke er noen lærebok i naturvitenskap, men spørsmålet om mennesket avstamning og tilknytning til Gud er for viktig til å overlate til de med samme syn som naturalismen i så måte. At Bibelen også henviser til den allmenne åpenbaring, er noe som kristen-darwinister ønsker vektlegge mindre. De anklager oss for ei 'samrøre' av tro og vitenskap. Men det er aldri vanntette skott mellom dem. Selv blir de gjerne mer opptatt av det politisk og vitenskapelige korrekte syn på jorda, og det dennesidige menneskeliv som det viktigste. Her og nå er det jo også det viktigste vi holder på med, men perspektivet er kortsiktig: Bibelen har ikke bare ord om en skapelse, men også om en nyskapelse. Det er utover perspektivet til en naturalistisk darwinisme. Uten å kunne dømme om dette, kan det vel synes som det ligger utenpå det mange kristen-darwinister synes er mest presserende.
At kristen-darwinistene omfavnes på en annen måte enn tilhengere av Intelligent Design, gjør nok sitt til at kristen-darwinister gjerne også tror seg bedre i stand til å hevde og forsvare kristne interesser overfor ateister. Noen avisredaksjoner synes også være av den oppfatning at det er nok at kristen-darwinister slipper til. Noen av dem sukrer da den bitre pillen ved å si at det er til gagn for kristenheten at de fører debatten for oss. Og det til tross for at de står omtrent for det motsatte av oss i denne saken. Det eneste positive jeg er i stand til å se, er at motsetninger kan skape en spenning og en debatt som gjør spørsmål om Guds rolle i verden mer sentral. Det vil jo ikke være en liten betydning, og det ville faktisk være verdt prisen å måtte stille i annen rekke ved diskusjoner etc.Kristen-darwinister hevder gjerne at vi står for en Bibelforståelse, som ikke griper Bibelens intensjoner. Det er jo lett å se for seg at Bibelen ikke er noen lærebok i naturvitenskap, men spørsmålet om mennesket avstamning og tilknytning til Gud er for viktig til å overlate til de med samme syn som naturalismen i så måte. At Bibelen også henviser til den allmenne åpenbaring, er noe som kristen-darwinister ønsker vektlegge mindre. De anklager oss for ei 'samrøre' av tro og vitenskap. Men det er aldri vanntette skott mellom dem. Selv blir de gjerne mer opptatt av det politisk og vitenskapelige korrekte syn på jorda, og det dennesidige menneskeliv som det viktigste. Her og nå er det jo også det viktigste vi holder på med, men perspektivet er kortsiktig: Bibelen har ikke bare ord om en skapelse, men også om en nyskapelse. Det er utover perspektivet til en naturalistisk darwinisme. Uten å kunne dømme om dette, kan det vel synes som det ligger utenpå det mange kristen-darwinister synes er mest presserende. i) Universitetsbiologene krever høre en bibelske begrunnelse fra oss, for at de skal kunne bevise vår manglende vitenskapelighet.
i) Universitetsbiologene krever høre en bibelske begrunnelse fra oss, for at de skal kunne bevise vår manglende vitenskapelighet. i) Den gir mennesket større frihet, intellektuelt sett. En er ikke bundet til å tro på kun dennesidig opphav til liv og artenes utvikling.
i) Den gir mennesket større frihet, intellektuelt sett. En er ikke bundet til å tro på kun dennesidig opphav til liv og artenes utvikling.
 Det er velkjent at den første Bibelske skapelsesberetning er lagt inn i et sjudagers skjema. Det må likevel hevdes at fra vitenskapelig hold kan det ikke være snakk om vanlige døgn, før jord og sol kom til. De dukker først opp den fjerde 'dagen'. Om en samstiller ord og vitenskap om relativistisk tidsforståelse, så virker det klart at det ikke er ment å forstå bokstavelig som syv døgn. Høydepunktet i beretningen er likevel at Gud skapte mennesket, i sitt bilde. Det innebærer at Gud står bak det, og at det har sin verdi i forhold til Ham. Vi har større grunn til å føle stolthet av vårt opphav enn darwinister har. Når det så gjelder varigheten av mennesker på jorda, har vi ikke noe fasitsvar på det.
Det er velkjent at den første Bibelske skapelsesberetning er lagt inn i et sjudagers skjema. Det må likevel hevdes at fra vitenskapelig hold kan det ikke være snakk om vanlige døgn, før jord og sol kom til. De dukker først opp den fjerde 'dagen'. Om en samstiller ord og vitenskap om relativistisk tidsforståelse, så virker det klart at det ikke er ment å forstå bokstavelig som syv døgn. Høydepunktet i beretningen er likevel at Gud skapte mennesket, i sitt bilde. Det innebærer at Gud står bak det, og at det har sin verdi i forhold til Ham. Vi har større grunn til å føle stolthet av vårt opphav enn darwinister har. Når det så gjelder varigheten av mennesker på jorda, har vi ikke noe fasitsvar på det. Om en legger sammen Bibelens år, har en bommet på det før -i forhold til vår tidsregnings begynnelse. At en kan bomme mer om en går helt tilbake til menneskets opprinnelse, skulle være mulig å forstå. Istider og andre katastrofer synes godt bevitnet i geologien. C-14 metoden synes godt dokumentert og begrunnet. Den har sin klare nullstilling av tiden, i forhold til hvordan andre 'radioaktive klokker blir nullstilt'. Men vi har også bemerket darwinisters ønske og behov for å skyve det meste inn i den utydelige, tåkebelagte fortid. At mennesket skulle framstå mer eller mindre på vårt nivå, uten muntlig og skriftlig språk i 90.000 år, og bare brølt og slått hverandre i hodet med steinkøller, sier mer om de som hevder det enn om hulemenneskene. Ut fra dette ville det bare dukke opp et nytt ord hvert tiår. Et et steinalderspråk på 9000 ord er relativt rikt. Ikke særlig kreativt må det sies. Ennå mer usannsynlig blir historien, slik noen forteller den: at mennesker i hovedsak har hatt sitt nåværende form og utseende mellom 100.000 til 1 million år før vår tisdregning..
Om en legger sammen Bibelens år, har en bommet på det før -i forhold til vår tidsregnings begynnelse. At en kan bomme mer om en går helt tilbake til menneskets opprinnelse, skulle være mulig å forstå. Istider og andre katastrofer synes godt bevitnet i geologien. C-14 metoden synes godt dokumentert og begrunnet. Den har sin klare nullstilling av tiden, i forhold til hvordan andre 'radioaktive klokker blir nullstilt'. Men vi har også bemerket darwinisters ønske og behov for å skyve det meste inn i den utydelige, tåkebelagte fortid. At mennesket skulle framstå mer eller mindre på vårt nivå, uten muntlig og skriftlig språk i 90.000 år, og bare brølt og slått hverandre i hodet med steinkøller, sier mer om de som hevder det enn om hulemenneskene. Ut fra dette ville det bare dukke opp et nytt ord hvert tiår. Et et steinalderspråk på 9000 ord er relativt rikt. Ikke særlig kreativt må det sies. Ennå mer usannsynlig blir historien, slik noen forteller den: at mennesker i hovedsak har hatt sitt nåværende form og utseende mellom 100.000 til 1 million år før vår tisdregning..
fritt etter 'Darwin 200 år -en festbrems' kap 1, av Peder A. Tyvand
 Darwinismens stilling i norske medier kan karakteriseres kortfattet: altoverveiende ene-dominerende. Enten det er natur eller kosmos, er det herskende holdning i NRK og andre hovedkanaler. Godt da at de må ty til innkjøp utenfra. Serien Gud og vitenskapen (januar 2014) ser i utgangspunktet ut til å slippe til ulike syn. I norskproduserte program får evolusjonister fritt boltre seg med sine lettvinte påstander om livets opphav og muligheter for liv i rommet etc. Det er sjelden noen i det hele får komme til motmæle. Mer typisk er det at evolusjonister hamrer løs mot karikerte, innbilte motstandere, uten mulighet for gjensvar. Darwinister har et knugende grep om undervisning, forskningsinstitusjoner og media. Maktposisjonen består, ikke minst fordi kritikk er sensurert vekk i over et halvt århundre. Vil man ikke vite av vitenskapelig kvalifisert kritikk av darwinismen? En hører ofte at kritikken kommer fra en håndfull 'kreasjonister' i USA. Men over 700 vitenskapsmenn fra hele verden med doktorgrad i naturvitenskap stiller sine navn bak kritikken. Alle disse har undertegnet et felles opprop med krav om at darwinismen utsettes for kritisk prøving. I Norge finnes det veldig få universitetsbiologer som tør risikere dette på grunn av faren for represalier.
Darwinismens stilling i norske medier kan karakteriseres kortfattet: altoverveiende ene-dominerende. Enten det er natur eller kosmos, er det herskende holdning i NRK og andre hovedkanaler. Godt da at de må ty til innkjøp utenfra. Serien Gud og vitenskapen (januar 2014) ser i utgangspunktet ut til å slippe til ulike syn. I norskproduserte program får evolusjonister fritt boltre seg med sine lettvinte påstander om livets opphav og muligheter for liv i rommet etc. Det er sjelden noen i det hele får komme til motmæle. Mer typisk er det at evolusjonister hamrer løs mot karikerte, innbilte motstandere, uten mulighet for gjensvar. Darwinister har et knugende grep om undervisning, forskningsinstitusjoner og media. Maktposisjonen består, ikke minst fordi kritikk er sensurert vekk i over et halvt århundre. Vil man ikke vite av vitenskapelig kvalifisert kritikk av darwinismen? En hører ofte at kritikken kommer fra en håndfull 'kreasjonister' i USA. Men over 700 vitenskapsmenn fra hele verden med doktorgrad i naturvitenskap stiller sine navn bak kritikken. Alle disse har undertegnet et felles opprop med krav om at darwinismen utsettes for kritisk prøving. I Norge finnes det veldig få universitetsbiologer som tør risikere dette på grunn av faren for represalier. Darwinismen er en mytebasert ideologi fra midten av 1800-tallet. Vi må ikke la oss dupere til å tro at den representerer fornuften. Fornuften viser seg alltid ved romslighet og åpenhet for debatt. Ufornuften og darwinismen gjør det stikk motsatte. Darwinismen er mer opptatt av makt enn fornuft. Det er viktigere å få rett enn å ha rett. Over 30 år som kritiker av darwinismen, har vist at det sjelden eller aldri åpnes for fri debatt. Når den skal konfrontere motstandere, er det ingen åpen, vitenskapelig tilnærming. En snakker om noen, i stedet for med noen. Det kalles på godt norsk for baktaling, ikke noe honnørord akkurat.
Darwinismen er en mytebasert ideologi fra midten av 1800-tallet. Vi må ikke la oss dupere til å tro at den representerer fornuften. Fornuften viser seg alltid ved romslighet og åpenhet for debatt. Ufornuften og darwinismen gjør det stikk motsatte. Darwinismen er mer opptatt av makt enn fornuft. Det er viktigere å få rett enn å ha rett. Over 30 år som kritiker av darwinismen, har vist at det sjelden eller aldri åpnes for fri debatt. Når den skal konfrontere motstandere, er det ingen åpen, vitenskapelig tilnærming. En snakker om noen, i stedet for med noen. Det kalles på godt norsk for baktaling, ikke noe honnørord akkurat.
 Om en kommer over argumentasjon fra darwinistisk hold, er det ofte en helt annen énsidig holdning: Påstand: Mennesket stammer fra en primitiv urcelle. Til støtte for dette har en to argumenter: DNA-basert genetikk og fossilfunn. En hevder at alt DNA og alle fossilfunn henger kontinuerlig sammen. En slår fast som et absolutt faktum at mennesker stammer fra en primitiv urcelle.
Om en kommer over argumentasjon fra darwinistisk hold, er det ofte en helt annen énsidig holdning: Påstand: Mennesket stammer fra en primitiv urcelle. Til støtte for dette har en to argumenter: DNA-basert genetikk og fossilfunn. En hevder at alt DNA og alle fossilfunn henger kontinuerlig sammen. En slår fast som et absolutt faktum at mennesker stammer fra en primitiv urcelle. I de senere år er det imidlertid kommet et nytt grunnlag innen faget med biologiske algoritmer. Algoritmene er kodet og lagret i DNA, og er hovedtemaet i denne boken. Til tross for at ledende darwinister vet at framtidens biologi vil bli en algoritmisk vitenskap, er det foreløpig få biologer som i noen grad har tatt det inn over seg. De som har arbeidet med algoritmer (matematikere/statistikere og datafolk) vet at de er sårbare for små tilfeldige endringer. Det er en merkelig og udokumentert påstand at en slår fast eksistensen av en sammenhengende rekke av funksjonelle, genetiske algoritmer fra urcelle til menneske. Spørsmålet er om alle ledd i denne rekken kunne dane funksjonelle koder for levende organismer.
I de senere år er det imidlertid kommet et nytt grunnlag innen faget med biologiske algoritmer. Algoritmene er kodet og lagret i DNA, og er hovedtemaet i denne boken. Til tross for at ledende darwinister vet at framtidens biologi vil bli en algoritmisk vitenskap, er det foreløpig få biologer som i noen grad har tatt det inn over seg. De som har arbeidet med algoritmer (matematikere/statistikere og datafolk) vet at de er sårbare for små tilfeldige endringer. Det er en merkelig og udokumentert påstand at en slår fast eksistensen av en sammenhengende rekke av funksjonelle, genetiske algoritmer fra urcelle til menneske. Spørsmålet er om alle ledd i denne rekken kunne dane funksjonelle koder for levende organismer. Konkret manifesterer problemet seg i form av forplantningsbarrierer. Hver gang det viser seg at en kryssbefruktning (hybridisering) mellom to gitte arter er umulig, setter det en effektiv stopper for kontinuiteten fra art til art. Det skyldes jo at DNA-kodingen har form som et dataprogram. Nye programdeler oppstår ikke av seg selv, i små trinn. Ingen har noensinne sett en lang kontinuerlig rekke av nesten like og fullt ut funksjonelle programmet. Dermed blir det brede og uoverstigelige barrierer mellom krypdyr, fugl og pattedyr. En kan ikke nekte folk å tro på en slik sammenhengende/kontinuerlig rekke av overganger. Men det krever stor tro å gjøre det! Det er verken empirisk eller fornuftsbasert, og alt bør ikke skje via skattebetalernes penger.
Konkret manifesterer problemet seg i form av forplantningsbarrierer. Hver gang det viser seg at en kryssbefruktning (hybridisering) mellom to gitte arter er umulig, setter det en effektiv stopper for kontinuiteten fra art til art. Det skyldes jo at DNA-kodingen har form som et dataprogram. Nye programdeler oppstår ikke av seg selv, i små trinn. Ingen har noensinne sett en lang kontinuerlig rekke av nesten like og fullt ut funksjonelle programmet. Dermed blir det brede og uoverstigelige barrierer mellom krypdyr, fugl og pattedyr. En kan ikke nekte folk å tro på en slik sammenhengende/kontinuerlig rekke av overganger. Men det krever stor tro å gjøre det! Det er verken empirisk eller fornuftsbasert, og alt bør ikke skje via skattebetalernes penger. sunn fornuft". Denne seieren har hatt en pris i form av filtrering, sensur og maktspråk gjennom minst 50 år i Norge.
sunn fornuft". Denne seieren har hatt en pris i form av filtrering, sensur og maktspråk gjennom minst 50 år i Norge. Det positive bidraget til Darwin er at han påpekte en viktig prosess som skjer innenfor en artspopulasjon, i et miljø hvor det er begrensede ressurser til rådighet. Da vil det naturlig utvalg (seleksjon) favorisere individer som er best tilpasset miljøet på bekostning av de individer som er dårligere tilpasset. Denne favoriseringen kan gå på at en lettere får tak i mat, kan unnslippe eller forsvare seg mot fiender. Sist men ikke minst at en lettere kan formere seg og bringe gunstige egenskaper videre til neste generasjon. De egenskapene som gjorde at en var bedre tilpasset et bestemt miljø enn de som ikke besatt disse egenskapene.
Det positive bidraget til Darwin er at han påpekte en viktig prosess som skjer innenfor en artspopulasjon, i et miljø hvor det er begrensede ressurser til rådighet. Da vil det naturlig utvalg (seleksjon) favorisere individer som er best tilpasset miljøet på bekostning av de individer som er dårligere tilpasset. Denne favoriseringen kan gå på at en lettere får tak i mat, kan unnslippe eller forsvare seg mot fiender. Sist men ikke minst at en lettere kan formere seg og bringe gunstige egenskaper videre til neste generasjon. De egenskapene som gjorde at en var bedre tilpasset et bestemt miljø enn de som ikke besatt disse egenskapene. På Darwins tid var det vanlig å tro at larver og annet småkryp oppsto av seg selv inne i en gjødselhaug, som fikk stå og 'godgjøre seg'. Denne fordommen hadde høy status og fikk sin egen vitenskapelige betegnelse: Spontan generasjon. Det er troen på at liv blir til av seg selv, når forholdene ligger til rette for det. Det innebærer at det finnes tilstrekkelig med næring, passende temperaturs og en passe dose med 'bakgrunnsstøy'. Darwins samtidige skulle der spille en større rolle enn han selv. Louis Pasteur (1822-1995) fant at liv bare kan bli til av liv. Pasteur gjorde mange eksperimenter der han foretok desinfisering ved kraftig varmebehandling slik at alle egg, frø og larver ble avlivet. Da fant han ut at inne i en fullstendig desinfisert gjødselhaug, ville det ikke komme et eneste lite kryp til verden. Ingenting kunne bli til i en gjødselhaug, om det ikke fantes livsspirer der allerede. Livsspirer som var kommet fra annet liv.
På Darwins tid var det vanlig å tro at larver og annet småkryp oppsto av seg selv inne i en gjødselhaug, som fikk stå og 'godgjøre seg'. Denne fordommen hadde høy status og fikk sin egen vitenskapelige betegnelse: Spontan generasjon. Det er troen på at liv blir til av seg selv, når forholdene ligger til rette for det. Det innebærer at det finnes tilstrekkelig med næring, passende temperaturs og en passe dose med 'bakgrunnsstøy'. Darwins samtidige skulle der spille en større rolle enn han selv. Louis Pasteur (1822-1995) fant at liv bare kan bli til av liv. Pasteur gjorde mange eksperimenter der han foretok desinfisering ved kraftig varmebehandling slik at alle egg, frø og larver ble avlivet. Da fant han ut at inne i en fullstendig desinfisert gjødselhaug, ville det ikke komme et eneste lite kryp til verden. Ingenting kunne bli til i en gjødselhaug, om det ikke fantes livsspirer der allerede. Livsspirer som var kommet fra annet liv.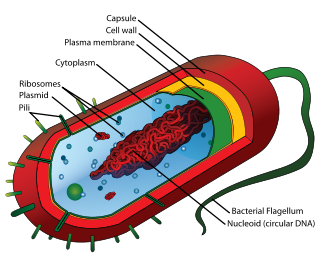 Pasteurs avlivning av spontan generasjon var et skudd for baugen på darwinismen, innerst inne har en aldri kunnet akseptere det nederlaget. Darwinismen må jo forutsette at alt liv blir til av seg selv, uten styring ovenfra. At livet skal ha en kilde høyere enn livet, kan den ikke tåle. Den har på forhånd vedtatt at livets kilde må ligge på et lavere plan enn livet. Møtet med Pasteurs overbevisende empirisme, førte til en strategiske tilbaketrekning. En flyttet påstanden om spontan generasjon langt bakover i tid, og nylig også utover i rommet. Noen hevder nå at brennende kometer bragte med seg materie i form av bakterier eller aminosyrer, som så er blitt til liv. Som om en bakterie uten kjerne, kan formere seg uten vert og næring, og at en brennende komet ikke ble like varm som Pasteurs gjødselhaug?
Pasteurs avlivning av spontan generasjon var et skudd for baugen på darwinismen, innerst inne har en aldri kunnet akseptere det nederlaget. Darwinismen må jo forutsette at alt liv blir til av seg selv, uten styring ovenfra. At livet skal ha en kilde høyere enn livet, kan den ikke tåle. Den har på forhånd vedtatt at livets kilde må ligge på et lavere plan enn livet. Møtet med Pasteurs overbevisende empirisme, førte til en strategiske tilbaketrekning. En flyttet påstanden om spontan generasjon langt bakover i tid, og nylig også utover i rommet. Noen hevder nå at brennende kometer bragte med seg materie i form av bakterier eller aminosyrer, som så er blitt til liv. Som om en bakterie uten kjerne, kan formere seg uten vert og næring, og at en brennende komet ikke ble like varm som Pasteurs gjødselhaug? Darwinismen hevder spontan-generasjon kun i ursuppen. Det var bare en fjern og enslig liten første 'nesten-celle', som ble til 'av seg selv'. Men nesten-cellen måtte ha noe å dannes ut fra. Så en tror proteiner ble dannet ved elektriske gnister i en ursuppe med enklere karbonforbindelser. Og så skulle disse proteinene videre ha organisert seg selv, laget sine egne cellemembraner og kjerner. Laget sine egne algoritmer, egne koder, egne avlesnings og reproduksjonsmekanismer. Dette spinkle nesten-liv var enten omgitt av oksygen, som en livsfarlig fiende. Eller omgitt av en atmosfære uten oksygen, slik at ånding for energiomsetning ikke var mulig. På toppen av dette, måtte dette første 'nesten-livet' lete etter næring, fordi enhver form for næring forutsetter liv. Intet liv kan overleve på uorganisk materiale (tygge stein). Ingen næringstilførsel kan trylles fram uten liv. Liv som må være i stand til å iverksette fotosyntese. I darwinistenes 'teorier' for livets opphav, er det med å skaffe seg næring helt ute av bildet. Det skal sterk tro til for å satse på dette, men 'må man så må man' virker det som.
Darwinismen hevder spontan-generasjon kun i ursuppen. Det var bare en fjern og enslig liten første 'nesten-celle', som ble til 'av seg selv'. Men nesten-cellen måtte ha noe å dannes ut fra. Så en tror proteiner ble dannet ved elektriske gnister i en ursuppe med enklere karbonforbindelser. Og så skulle disse proteinene videre ha organisert seg selv, laget sine egne cellemembraner og kjerner. Laget sine egne algoritmer, egne koder, egne avlesnings og reproduksjonsmekanismer. Dette spinkle nesten-liv var enten omgitt av oksygen, som en livsfarlig fiende. Eller omgitt av en atmosfære uten oksygen, slik at ånding for energiomsetning ikke var mulig. På toppen av dette, måtte dette første 'nesten-livet' lete etter næring, fordi enhver form for næring forutsetter liv. Intet liv kan overleve på uorganisk materiale (tygge stein). Ingen næringstilførsel kan trylles fram uten liv. Liv som må være i stand til å iverksette fotosyntese. I darwinistenes 'teorier' for livets opphav, er det med å skaffe seg næring helt ute av bildet. Det skal sterk tro til for å satse på dette, men 'må man så må man' virker det som. Det er ikke bare overtroen på spontan generasjon som er flyttet bakover i tid. Det samme unnvikelsesmanøveren brukes for alle betente overganger mellom artene på vei fra amøbe til menneske. Det er visstnok ingen eksisterende dyr som stammer fra andre typer konkrete dyr lenger. Mennesket stammer ikke lenger fra apene, men 'har felles apelignende forfedre med apene.' En flytter hele spekulasjonen om en felles avstamning akkurat passe langt tilbake i fortidas tåkeheim, til at det skal være umulig å motsi spekulasjonen. Darwinismen er gjennomsyret av en grunnleggende lettvinthet i forhold til hvor lett det er å få til liv. Utdøing er ikke noe problem, det gir bare ledige økologiske nisjer.
Det er ikke bare overtroen på spontan generasjon som er flyttet bakover i tid. Det samme unnvikelsesmanøveren brukes for alle betente overganger mellom artene på vei fra amøbe til menneske. Det er visstnok ingen eksisterende dyr som stammer fra andre typer konkrete dyr lenger. Mennesket stammer ikke lenger fra apene, men 'har felles apelignende forfedre med apene.' En flytter hele spekulasjonen om en felles avstamning akkurat passe langt tilbake i fortidas tåkeheim, til at det skal være umulig å motsi spekulasjonen. Darwinismen er gjennomsyret av en grunnleggende lettvinthet i forhold til hvor lett det er å få til liv. Utdøing er ikke noe problem, det gir bare ledige økologiske nisjer. Dagens opplyste mennesker mener å kjenne til DNA-koden. Men DNA i seg selv er ingen kilde til informasjon, eller noen kode. DNA er bærer av informasjon, og det er noe annet. Informasjonen hos DNA ligger ikke i selve dobbeltspiralen, men kun i trappetrinnene som holder de to enkeltspiralene sammen, slik at de blir en dobbeltspiral. Kodingskonvensjonen ligger ikke internt i DNA-spiralen. Den må være gitt på forhånd, eksternt. Vi skal se at darwinister forkvakler informasjonsbegrepet. Det skjer bl.a. ved at informasjonskilde og informasjonsbærer rotes i hop. De fleste mennesker har imidlertid en naturlig forståelse av dette: Energikilder er stoffer som gir energi: kull, olje, gass og fossefall. Energibærere er medier som i utgangspunktet ikke har energi i seg, men må få den utenfra, slik som høyspentledninger og oppladbare batterier etc. At hydrogen framføres som energikilde av noen, har den haken at det ikke finnes i fri form på jorda. Da kan det kreve ganske mye energi å utvinne det.
Dagens opplyste mennesker mener å kjenne til DNA-koden. Men DNA i seg selv er ingen kilde til informasjon, eller noen kode. DNA er bærer av informasjon, og det er noe annet. Informasjonen hos DNA ligger ikke i selve dobbeltspiralen, men kun i trappetrinnene som holder de to enkeltspiralene sammen, slik at de blir en dobbeltspiral. Kodingskonvensjonen ligger ikke internt i DNA-spiralen. Den må være gitt på forhånd, eksternt. Vi skal se at darwinister forkvakler informasjonsbegrepet. Det skjer bl.a. ved at informasjonskilde og informasjonsbærer rotes i hop. De fleste mennesker har imidlertid en naturlig forståelse av dette: Energikilder er stoffer som gir energi: kull, olje, gass og fossefall. Energibærere er medier som i utgangspunktet ikke har energi i seg, men må få den utenfra, slik som høyspentledninger og oppladbare batterier etc. At hydrogen framføres som energikilde av noen, har den haken at det ikke finnes i fri form på jorda. Da kan det kreve ganske mye energi å utvinne det. Informasjon er et oversanselig (metafysisk) begrep, fordi det alltid inneholder viktige ikke-sansbare elementer. Et grunnleggende bilde av informasjonsoverføring går som følger:Noen av oss lærte som barn at 'alt det gode kommer ovenfra'. Grunntanken i darwinismen er motsatt: 'Alle årsaker kommer nedenfra'. I darwinistisk tankegang er ikke livet noe som skjenkes oss ovenfra, tvert i mot kommer det fra pølen der nede. Livet kommer snikende fra en uidentifiserbar amøbe, nede i 'ursuppa'. Denne amøben har dratt seg opp etter håret den ikke har, og er blitt mer og mer avansert på sin vei til å bli menneske. Det må være et filosofisk og logisk problem for darwinismen at alle forklaringer er fattigere enn fenomenene de skal forklare. Ved stadig reduksjon blir uvegerlig noe borte underveis. Det at liv spontant skal ha blitt til av ikke-liv er et slikt eksempel.
Informasjon er et oversanselig (metafysisk) begrep, fordi det alltid inneholder viktige ikke-sansbare elementer. Et grunnleggende bilde av informasjonsoverføring går som følger:Noen av oss lærte som barn at 'alt det gode kommer ovenfra'. Grunntanken i darwinismen er motsatt: 'Alle årsaker kommer nedenfra'. I darwinistisk tankegang er ikke livet noe som skjenkes oss ovenfra, tvert i mot kommer det fra pølen der nede. Livet kommer snikende fra en uidentifiserbar amøbe, nede i 'ursuppa'. Denne amøben har dratt seg opp etter håret den ikke har, og er blitt mer og mer avansert på sin vei til å bli menneske. Det må være et filosofisk og logisk problem for darwinismen at alle forklaringer er fattigere enn fenomenene de skal forklare. Ved stadig reduksjon blir uvegerlig noe borte underveis. Det at liv spontant skal ha blitt til av ikke-liv er et slikt eksempel. Richard Dawkins som har vunnet enorm popularitet, bl.a. ved sin bok 'The selfish gene' uttrykker der en naiv overtro på det naturlige utvalg som årsak nedenfra. Det kan sammenlignes med at han oppfører seg som et milliardær-enebarn med bare fine leker. Et bortskjemt barn som er overbevist om at han har bare fine leker fordi kaster alle som ikke er det. For et barn kan kanskje kasting være fullgod forklaring på at det gjenværende er fint. Det har den fordelen for barnet at det er takket være sin egen kasting at alle hans leker er fine. Han trenger bare takke seg selv som er så klok at han systematisk kaster alt som ikke er helt topp.
Richard Dawkins som har vunnet enorm popularitet, bl.a. ved sin bok 'The selfish gene' uttrykker der en naiv overtro på det naturlige utvalg som årsak nedenfra. Det kan sammenlignes med at han oppfører seg som et milliardær-enebarn med bare fine leker. Et bortskjemt barn som er overbevist om at han har bare fine leker fordi kaster alle som ikke er det. For et barn kan kanskje kasting være fullgod forklaring på at det gjenværende er fint. Det har den fordelen for barnet at det er takket være sin egen kasting at alle hans leker er fine. Han trenger bare takke seg selv som er så klok at han systematisk kaster alt som ikke er helt topp. Men hva skal en eller hjelpe seg med? Når en står overfor livet i dets rikdom og fylde, og eneste middel en har for å forstå det er reduksjon. En er pent nødt til å innbille seg at en forstår livet ved å kassere ting. Men om en reduserer livet, er det ikke livet en forstår. En bare fjerner og tabubelegger de virkelige årsakene. Og en bidrar til å fjerne essensen i informasjonsbegrepet. En må nesten forsøke redusere informasjonsbegrepet til noe materielt, om en vil hindre at noen årsak kan komme ovenfra. Konsekvensen blir at en bryter ned allmuens sunne fornuft under dekke av å være visere enn allmuen. Det får også den rare konsekvensen at resultatet kan bli bedre enn forutsetningene.
Men hva skal en eller hjelpe seg med? Når en står overfor livet i dets rikdom og fylde, og eneste middel en har for å forstå det er reduksjon. En er pent nødt til å innbille seg at en forstår livet ved å kassere ting. Men om en reduserer livet, er det ikke livet en forstår. En bare fjerner og tabubelegger de virkelige årsakene. Og en bidrar til å fjerne essensen i informasjonsbegrepet. En må nesten forsøke redusere informasjonsbegrepet til noe materielt, om en vil hindre at noen årsak kan komme ovenfra. Konsekvensen blir at en bryter ned allmuens sunne fornuft under dekke av å være visere enn allmuen. Det får også den rare konsekvensen at resultatet kan bli bedre enn forutsetningene. For kunst og musikk kan det medvirke til at en tror at verket er rikere enn kunstnerens sinn, eller rikere enn fargesammensetningene til rådighet på paletten. En kan innbille seg at komposisjonen holder et høyere musikalsk nivå enn komponisten. En tror maleriet blir vakrere enn forestillingen kunstneren hadde i sitt hode, da han laget det. Denne tankegangen har et filosofisk forklaringspoblem: Noe framstilles uten fullverdig årsak, -hvor kommer 'resten' fra? Men dette materialistiske prosjektet er ikke så viselig at det lar seg gjennomføre konsekvent.
For kunst og musikk kan det medvirke til at en tror at verket er rikere enn kunstnerens sinn, eller rikere enn fargesammensetningene til rådighet på paletten. En kan innbille seg at komposisjonen holder et høyere musikalsk nivå enn komponisten. En tror maleriet blir vakrere enn forestillingen kunstneren hadde i sitt hode, da han laget det. Denne tankegangen har et filosofisk forklaringspoblem: Noe framstilles uten fullverdig årsak, -hvor kommer 'resten' fra? Men dette materialistiske prosjektet er ikke så viselig at det lar seg gjennomføre konsekvent. Det materialistiske synet på lov og rett, går under navnet rettspositivisme. En rettspositivist vil se på en forbrytelse som et resultat av den arv og det miljø forbryteren springer ut fra og bærer med seg. Dette grunnsynet vil så langt mulig hente alle årsakssammenhenger nedenfra: fra en viljeløs arv og et ansvarsfritt miljø, som den stakkars forbryteren er prisgitt. En rettspositivistisk etterforskning vil gjerne vektlegge gjerningsmannens ansvar og motiv minimalt. Når det gjelder forbryteren, har en rettspositivist intet ønske om en årsak ovenfra. Når det gjelder offeret, er det naturligvis beklagelig, men gjort er gjort og 'det vil ikke hjelpe offeret om forbryteren straffes hardere'.. På papiret er saken forstått til bunns, på samme måte som en darwinist forstår livet til bunns, I sine egne tankesirkler vel å merke.
Det materialistiske synet på lov og rett, går under navnet rettspositivisme. En rettspositivist vil se på en forbrytelse som et resultat av den arv og det miljø forbryteren springer ut fra og bærer med seg. Dette grunnsynet vil så langt mulig hente alle årsakssammenhenger nedenfra: fra en viljeløs arv og et ansvarsfritt miljø, som den stakkars forbryteren er prisgitt. En rettspositivistisk etterforskning vil gjerne vektlegge gjerningsmannens ansvar og motiv minimalt. Når det gjelder forbryteren, har en rettspositivist intet ønske om en årsak ovenfra. Når det gjelder offeret, er det naturligvis beklagelig, men gjort er gjort og 'det vil ikke hjelpe offeret om forbryteren straffes hardere'.. På papiret er saken forstått til bunns, på samme måte som en darwinist forstår livet til bunns, I sine egne tankesirkler vel å merke. Men når en rettspositivist kommer til rettsalen, til virkeligheten så og si, må de også forholde seg til motiv for handlingen. Årsaken til forbrytelser må hentes ovenfra, fra forbryterens eget sinn og vilje. Å frita mennesket for ansvar for sine handlinger, er å degradere det til noe mindre enn mennesker. Når det konfronteres med det virkelige liv, faller grublerier om at alle årsaker skyldes arv og miljø til jorden. Men darwinister gjør akkurat samme feil, når de uten grunn, dogmatisk slår fast at alle årsaker er primitive og kommer nedenfra. Det er ikke livets årsaker, men darwinismens oppfatninger om dem som er primitive. Darwinisme handler om å projisere sine forestillinger over på livet og kalle denne projeksjonen en forklaring.Biologiprofessorene Stenseth og Høiland skrev i Aftenposten 13.okt. 2008: 'Charles Darwin viser at vi mennesker er en del av naturen som omgir oss. Nå forstår vi menneskets opprinnelse.' Dette utsagnet er litt drøyt, særlig når det er to sprenglærde professorer som uttaler seg. De forsøker antagelig fortelle at det naturlige utvalg er årsak til menneskets opprinnelse. Innsikten som disse to har om det naturlige utvalg, gjør at de synes de forstår fullt ut hvordan mennesket liksom ble til av laverestående vesener. Men forståelsen til menneskets opprinnelse, blir også en forståelse av hodet til mennesket hvor denne forståelsen angivelig sitter. Det blir dermed en forståelse av forståelsen til 'forståeren'. Her er fristende å legge til 'forstå-seg-påeren'.
Men når en rettspositivist kommer til rettsalen, til virkeligheten så og si, må de også forholde seg til motiv for handlingen. Årsaken til forbrytelser må hentes ovenfra, fra forbryterens eget sinn og vilje. Å frita mennesket for ansvar for sine handlinger, er å degradere det til noe mindre enn mennesker. Når det konfronteres med det virkelige liv, faller grublerier om at alle årsaker skyldes arv og miljø til jorden. Men darwinister gjør akkurat samme feil, når de uten grunn, dogmatisk slår fast at alle årsaker er primitive og kommer nedenfra. Det er ikke livets årsaker, men darwinismens oppfatninger om dem som er primitive. Darwinisme handler om å projisere sine forestillinger over på livet og kalle denne projeksjonen en forklaring.Biologiprofessorene Stenseth og Høiland skrev i Aftenposten 13.okt. 2008: 'Charles Darwin viser at vi mennesker er en del av naturen som omgir oss. Nå forstår vi menneskets opprinnelse.' Dette utsagnet er litt drøyt, særlig når det er to sprenglærde professorer som uttaler seg. De forsøker antagelig fortelle at det naturlige utvalg er årsak til menneskets opprinnelse. Innsikten som disse to har om det naturlige utvalg, gjør at de synes de forstår fullt ut hvordan mennesket liksom ble til av laverestående vesener. Men forståelsen til menneskets opprinnelse, blir også en forståelse av hodet til mennesket hvor denne forståelsen angivelig sitter. Det blir dermed en forståelse av forståelsen til 'forståeren'. Her er fristende å legge til 'forstå-seg-påeren'. Det er lov å prøve seg. Men det å tro at en forstår noe er ikke ensbetydende med at en faktisk gjør det. Biologiprofessorene har rett i at det er mulig å forstå det naturlige utvalg. Det naturlige utvalg har bare et trikk til rådighet: ' Å velge bort ting.' Det naturlige utvalgs repertoar kan sammenlignes med trubaduren Paul Simons 'One Trick Pony'. Denne ponnien hadde kun ett trick til rådighet der den reiste rundt og viste seg fram. Dermed nådde den ikke den store populariteten, men det naturlige utvalg har blitt den store vinneren! Om det ikke hjelper oss å forstå naturen, så har det i hvert fall hjulpet darwinismen å bli vinneren så langt. Men det kan snu! Tricket til ponnien var sikkert mer avansert enn det naturlige utvalg kan varte opp med.
Det er lov å prøve seg. Men det å tro at en forstår noe er ikke ensbetydende med at en faktisk gjør det. Biologiprofessorene har rett i at det er mulig å forstå det naturlige utvalg. Det naturlige utvalg har bare et trikk til rådighet: ' Å velge bort ting.' Det naturlige utvalgs repertoar kan sammenlignes med trubaduren Paul Simons 'One Trick Pony'. Denne ponnien hadde kun ett trick til rådighet der den reiste rundt og viste seg fram. Dermed nådde den ikke den store populariteten, men det naturlige utvalg har blitt den store vinneren! Om det ikke hjelper oss å forstå naturen, så har det i hvert fall hjulpet darwinismen å bli vinneren så langt. Men det kan snu! Tricket til ponnien var sikkert mer avansert enn det naturlige utvalg kan varte opp med. Hansen, tidligere filmguru i NRK, der han innleder med å si. 'Nå skal vi presentere et utvalg av påskens nye filmer.' Han har valgt bort mange filmer, ut fra sine preferanser. Hensikten er å gi plass til de få som kan presenteres innenfor en begrenset tidsramme. Det er ikke slik at Pål Bang-Hansens utvalg er den egentlige årsak til de filmene som presenteres. Det at filmene var produsert og klare for visning var den bakenforliggende årsaken. Men det virker fortsatt som darwinister vil ha oss til å tro at: 'Naturlig utvalg er en dominerende faktor m.h.t. utforming av det biologiske livs mangfold,' slik professor Stig W. Omholt uttrykte det (Stenseth & Lie 1984, s177)
Hansen, tidligere filmguru i NRK, der han innleder med å si. 'Nå skal vi presentere et utvalg av påskens nye filmer.' Han har valgt bort mange filmer, ut fra sine preferanser. Hensikten er å gi plass til de få som kan presenteres innenfor en begrenset tidsramme. Det er ikke slik at Pål Bang-Hansens utvalg er den egentlige årsak til de filmene som presenteres. Det at filmene var produsert og klare for visning var den bakenforliggende årsaken. Men det virker fortsatt som darwinister vil ha oss til å tro at: 'Naturlig utvalg er en dominerende faktor m.h.t. utforming av det biologiske livs mangfold,' slik professor Stig W. Omholt uttrykte det (Stenseth & Lie 1984, s177) Omholt ønsket sikkert påpeke at naturlig utvalg er ansvarlig for at de genfrekvensene vi observerer i naturlige populasjoner er slik de er. Men det er ikke det samme som mangfold. Mangfoldet påvirkes negativt av det naturlige utvalg, ved at genetiske egenskaper dør ut fortere enn om seleksjonen ikke hadde vært virksom. Sitatet ovenfor hadde vært bedre tilpasset virkeligheten om ordet 'mangfold' hadde vært byttet ut med 'begrensning'. Det er heller slik at naturlig utvalg er mangfoldets fiende enn dets kilde. Dette er et eksempel på at i darwinismen er en fornøyd med overflatiske spekulasjoner, i stedet for å gå i dybden etter forklaringer. En skulle tro nye egenskaper ble utviklet på grunn av seleksjon, slik de snakker. Men naturlig utvalg virker alltid på egenskaper som allerede er tilstede.
Omholt ønsket sikkert påpeke at naturlig utvalg er ansvarlig for at de genfrekvensene vi observerer i naturlige populasjoner er slik de er. Men det er ikke det samme som mangfold. Mangfoldet påvirkes negativt av det naturlige utvalg, ved at genetiske egenskaper dør ut fortere enn om seleksjonen ikke hadde vært virksom. Sitatet ovenfor hadde vært bedre tilpasset virkeligheten om ordet 'mangfold' hadde vært byttet ut med 'begrensning'. Det er heller slik at naturlig utvalg er mangfoldets fiende enn dets kilde. Dette er et eksempel på at i darwinismen er en fornøyd med overflatiske spekulasjoner, i stedet for å gå i dybden etter forklaringer. En skulle tro nye egenskaper ble utviklet på grunn av seleksjon, slik de snakker. Men naturlig utvalg virker alltid på egenskaper som allerede er tilstede. Det er imidlertid en dypereliggende grunn til at det kan virke som nye egenskaper dukker opp via naturlig utvalg, og det er at mange genetiske egenskaper er recessive: "recessiv" kommer fra latin og betyr "vikende". Recessive gener er gener som er vikende i forhold til dominante gener. Dominante gener kommer til uttrykk om de er arvet bare fra en av de biologiske foreldrene. Recessivt virkende gener må arves fra begge foreldre for at de skal komme til uttrykk. Et velkjent eksempel er at gener for brune øyne er dominante, og at gener for blå øyne er recessive.
Det er imidlertid en dypereliggende grunn til at det kan virke som nye egenskaper dukker opp via naturlig utvalg, og det er at mange genetiske egenskaper er recessive: "recessiv" kommer fra latin og betyr "vikende". Recessive gener er gener som er vikende i forhold til dominante gener. Dominante gener kommer til uttrykk om de er arvet bare fra en av de biologiske foreldrene. Recessivt virkende gener må arves fra begge foreldre for at de skal komme til uttrykk. Et velkjent eksempel er at gener for brune øyne er dominante, og at gener for blå øyne er recessive. Vi og de fleste dyr og planter er diploide vesener. Det innebærer at arvestoffet vårt består av kromosompar. Hvert kromosompar har dobbel dose av genetisk koding, der den ene delen kommer fra far og den andre fra mor. Ofte er den genetiske informasjonen i det ene kromosomet i et par dominerende, slik at innholdet i det andre paret undertrykkes. Mange av våre mutasjonsskader er recessive. Dersom alle våre mutasjoner hadde kommet til uttrykk, hadde det ikke vært mange friske mennesker. Våre friske dominante gener redder oss fra våre syke recessive gener gang på gang. Men recessive gener kan også inneholde svært verdifull informasjon. Til tider kan det være livsviktig for en populasjon.
Vi og de fleste dyr og planter er diploide vesener. Det innebærer at arvestoffet vårt består av kromosompar. Hvert kromosompar har dobbel dose av genetisk koding, der den ene delen kommer fra far og den andre fra mor. Ofte er den genetiske informasjonen i det ene kromosomet i et par dominerende, slik at innholdet i det andre paret undertrykkes. Mange av våre mutasjonsskader er recessive. Dersom alle våre mutasjoner hadde kommet til uttrykk, hadde det ikke vært mange friske mennesker. Våre friske dominante gener redder oss fra våre syke recessive gener gang på gang. Men recessive gener kan også inneholde svært verdifull informasjon. Til tider kan det være livsviktig for en populasjon. ikke er gunstige i det endrede miljøet. Om de skjulte egenskapene ligger godt skjult, kan det ta tid for dem å bli virksomme. Dersom miljøendringene skjer raskere enn de skjulte egenskapene rekker å komme fram, holder ikke den genetiske driften tritt med seleksjonspresset. Da kan hele populasjoner dø ut. Dette skjedde i mange norske fiskevann på 70-tallet, grunnet sur nedbør. Det naturlige utvalg er en metode for å få gjort genetisk informasjon operativ. Det må ikke forveksles med en årsak. Kaldpressing er beste metode for å lage olivenolje med høy kvalitet. Om resultatet blir bedre enn varmebehandling, kan en lett tilskrive kaldpressingen som årsak til den bra kvaliteten. Men den tilførte ikke oljen noen egenskaper frukten ikke hadde i utgangspunktet. En slik overtro på en metode ligner den darwinister legger for dagen i forhold til naturlig utvalg.
ikke er gunstige i det endrede miljøet. Om de skjulte egenskapene ligger godt skjult, kan det ta tid for dem å bli virksomme. Dersom miljøendringene skjer raskere enn de skjulte egenskapene rekker å komme fram, holder ikke den genetiske driften tritt med seleksjonspresset. Da kan hele populasjoner dø ut. Dette skjedde i mange norske fiskevann på 70-tallet, grunnet sur nedbør. Det naturlige utvalg er en metode for å få gjort genetisk informasjon operativ. Det må ikke forveksles med en årsak. Kaldpressing er beste metode for å lage olivenolje med høy kvalitet. Om resultatet blir bedre enn varmebehandling, kan en lett tilskrive kaldpressingen som årsak til den bra kvaliteten. Men den tilførte ikke oljen noen egenskaper frukten ikke hadde i utgangspunktet. En slik overtro på en metode ligner den darwinister legger for dagen i forhold til naturlig utvalg. Å sammenligne naturlig utvalg med tilfeldig genetisk drift blir omtrent som å sammenligne en bred sakteflytende flod med en elv som går i et bratt fossestryk. Sammenlignet med fossefallet kan elven betraktes som en horisontal strømning. Men derav kan en ikke slutte at den strømmer svakt oppover! Tyngdekraften gjelder i virkelighetens verden. I den darwinistiske verden hevdes at mangfoldet øker på grunn av naturlig utvalg. Det er i beste fall sammenlignet med hva mangfoldet ville ha gjort ved tilfeldig genetisk drift. Dermed kan det med en viss rett hevdes at naturlig utvalg virker konserverende. Om en opplever darwinistisk lovprisning av seleksjon, kan en huske at om den ikke forklarer naturen, så forklarer den darwinismens hegemoni. Og det holder i massevis ser det ut til.
Å sammenligne naturlig utvalg med tilfeldig genetisk drift blir omtrent som å sammenligne en bred sakteflytende flod med en elv som går i et bratt fossestryk. Sammenlignet med fossefallet kan elven betraktes som en horisontal strømning. Men derav kan en ikke slutte at den strømmer svakt oppover! Tyngdekraften gjelder i virkelighetens verden. I den darwinistiske verden hevdes at mangfoldet øker på grunn av naturlig utvalg. Det er i beste fall sammenlignet med hva mangfoldet ville ha gjort ved tilfeldig genetisk drift. Dermed kan det med en viss rett hevdes at naturlig utvalg virker konserverende. Om en opplever darwinistisk lovprisning av seleksjon, kan en huske at om den ikke forklarer naturen, så forklarer den darwinismens hegemoni. Og det holder i massevis ser det ut til. Darwinister har laget sin egen seleksjonsliturgi der følgende trosbekjennelse framføres: 'Denne egenskapen (seleksjon) utviklet seg fordi det var gunstig for overlevelse'. I NRK's naturprogram på TV, ble ordet overlevelse på brukt på en spesiell måte: 'overlevelse i form av seier i konkurransen om økologiske nisjer'. I følge darwinismen må alltid seleksjonen være årsak til spesielle egenskaper hos en art. Men la oss se litt grundigere på dette. Det kan faktis finnes andre og flere muligheter: i) En annen og dypere årsak kan ligge bak det naturlige utvalg. ii) Det naturlige utvalg kan være en virkning i stedet for en årsak. iii) Det naturlige utvalg er verken en årsak eller en virkning.
Darwinister har laget sin egen seleksjonsliturgi der følgende trosbekjennelse framføres: 'Denne egenskapen (seleksjon) utviklet seg fordi det var gunstig for overlevelse'. I NRK's naturprogram på TV, ble ordet overlevelse på brukt på en spesiell måte: 'overlevelse i form av seier i konkurransen om økologiske nisjer'. I følge darwinismen må alltid seleksjonen være årsak til spesielle egenskaper hos en art. Men la oss se litt grundigere på dette. Det kan faktis finnes andre og flere muligheter: i) En annen og dypere årsak kan ligge bak det naturlige utvalg. ii) Det naturlige utvalg kan være en virkning i stedet for en årsak. iii) Det naturlige utvalg er verken en årsak eller en virkning. Vi så ovenfor at recessive gener ofte er den dypere og egentlige årsak bak fenomener som kan se ut som resultat av det naturlige utvalg. Vi har nevnt ørretens evne til å stå imot sur nedbør. Det naturlige utvalg er bare en metode for å lokke fram egenskaper som ligger godt gjemt i artens arvestoff. Det naturlige utvalg er heller ingen årsak til konkurransen om økologiske nisjer. Det er heller motsatt. Det er konkurransen om begrensede goder som er den bakenforliggende årsaken. Og det naturlige utvalg er virkningen av denne årsaken. Det naturlige utvalg er heller ikke årsak til livets opprinnelse. Der måtte det noe kvalitativt nytt til. For å fylle økologiske nisjer er det et annet prinsipp som gjelder: 'førstemann til mølla..' Det er først når konkurransen tetner seg til at det naturlige utvalg trår i kraft.
Vi så ovenfor at recessive gener ofte er den dypere og egentlige årsak bak fenomener som kan se ut som resultat av det naturlige utvalg. Vi har nevnt ørretens evne til å stå imot sur nedbør. Det naturlige utvalg er bare en metode for å lokke fram egenskaper som ligger godt gjemt i artens arvestoff. Det naturlige utvalg er heller ingen årsak til konkurransen om økologiske nisjer. Det er heller motsatt. Det er konkurransen om begrensede goder som er den bakenforliggende årsaken. Og det naturlige utvalg er virkningen av denne årsaken. Det naturlige utvalg er heller ikke årsak til livets opprinnelse. Der måtte det noe kvalitativt nytt til. For å fylle økologiske nisjer er det et annet prinsipp som gjelder: 'førstemann til mølla..' Det er først når konkurransen tetner seg til at det naturlige utvalg trår i kraft. Det naturlige utvalg er heller aldri årsak til overlevelse. Overlevelsen har høyere prioritet. Først da den grunnleggende overlevelse er på plass, kan det finnes noe å drive seleksjon på. Det må være liv, for å være håp! I følge darwinismen blir livet ofte nærmest tas for gitt. Er det ikke en livsform som benytter seg av mulighetene, blir det en annen. 'So what?' Det har imidlertid vært kriseperioder for enkelte populasjoner, og kan fort bli det igjen med svingninger i miljø og klimavariable. For noen år siden var det bibestanden som var utsatt. Mange viktige avlinger er avhengig av bestøvning fra bier. Ingen andre dyr bidrar så mye til planteproduksjon som det bier gjør. Pollinering (bestøvning) er livets motor, som sørger for at plantene kan leve videre. I følge en EU-publikasjon er 76% av matproduksjonen i verden avhengig av biavlen. I Europa er 84% av matproduksjonen avhengig av pollinering. Darwinismens bidrag i en slik krisetid, var deprimerende. Det kom mest etterpåkloke ord og påstander, lite av konkrete råd og virkemidler. Dersom bare noen få prosent av midlene brukt til mutasjoner på bananfluer var benyttet på bier, kunne en vært mye bedre rustet til å møte krisen. Bier er et sosialt insekt. Ansvarlig forfatter Peder A. Tyvand har i den sammenheng bidratt med en populasjonsgenetisk modell for sosiale insekter, som kan være til konkret hjelp for bestandens overlevelse.
Det naturlige utvalg er heller aldri årsak til overlevelse. Overlevelsen har høyere prioritet. Først da den grunnleggende overlevelse er på plass, kan det finnes noe å drive seleksjon på. Det må være liv, for å være håp! I følge darwinismen blir livet ofte nærmest tas for gitt. Er det ikke en livsform som benytter seg av mulighetene, blir det en annen. 'So what?' Det har imidlertid vært kriseperioder for enkelte populasjoner, og kan fort bli det igjen med svingninger i miljø og klimavariable. For noen år siden var det bibestanden som var utsatt. Mange viktige avlinger er avhengig av bestøvning fra bier. Ingen andre dyr bidrar så mye til planteproduksjon som det bier gjør. Pollinering (bestøvning) er livets motor, som sørger for at plantene kan leve videre. I følge en EU-publikasjon er 76% av matproduksjonen i verden avhengig av biavlen. I Europa er 84% av matproduksjonen avhengig av pollinering. Darwinismens bidrag i en slik krisetid, var deprimerende. Det kom mest etterpåkloke ord og påstander, lite av konkrete råd og virkemidler. Dersom bare noen få prosent av midlene brukt til mutasjoner på bananfluer var benyttet på bier, kunne en vært mye bedre rustet til å møte krisen. Bier er et sosialt insekt. Ansvarlig forfatter Peder A. Tyvand har i den sammenheng bidratt med en populasjonsgenetisk modell for sosiale insekter, som kan være til konkret hjelp for bestandens overlevelse. Så kan en alltids undres hvorfor darwinismen hadde så lite konkret å bidra med når våre viktigste husdyr var i krise. Etter vår mening er det fordi darwinismen er en tankeflukt. En akrobatisk øvelse i å tenke i strid med virkeligheten. Darwinismens såkalte mekanismer snur tingene fullstendig på hodet når det gjelder byggende kontra destruktive krefter i tilværelsen:
Så kan en alltids undres hvorfor darwinismen hadde så lite konkret å bidra med når våre viktigste husdyr var i krise. Etter vår mening er det fordi darwinismen er en tankeflukt. En akrobatisk øvelse i å tenke i strid med virkeligheten. Darwinismens såkalte mekanismer snur tingene fullstendig på hodet når det gjelder byggende kontra destruktive krefter i tilværelsen: Egentlig er det ganske hårreisende at det er mulig å gjøre disse negative kreftene positivt ansvarlig for alt liv som utfolder seg på planeten. I stedet for å reise bust mot denne pseudo-åndelige, kvasi-intellektuelle tankegangen, bøyer mange hodet uten å mukke under det destruktive åket. Teologer av god vilje hevder på ramme alvor at kirkens skapelses-formidling må være et lydrike under darwinismens mytologi. Og det er en mytologi på kollisjonskurs med realitetene. La oss se mer på hvordan darwinismens to 'mekanismer' blir tillagt egenskaper stikk motsatt av det de har:1) Tilfeldige mutasjoner får æren for oppbygningen av biologiens orden og genetiske informasjon. I virkeligheten virker mutasjonene ensidig og irreversibelt nedbrytende på biologisk orden og genetisk informasjon.
Egentlig er det ganske hårreisende at det er mulig å gjøre disse negative kreftene positivt ansvarlig for alt liv som utfolder seg på planeten. I stedet for å reise bust mot denne pseudo-åndelige, kvasi-intellektuelle tankegangen, bøyer mange hodet uten å mukke under det destruktive åket. Teologer av god vilje hevder på ramme alvor at kirkens skapelses-formidling må være et lydrike under darwinismens mytologi. Og det er en mytologi på kollisjonskurs med realitetene. La oss se mer på hvordan darwinismens to 'mekanismer' blir tillagt egenskaper stikk motsatt av det de har:1) Tilfeldige mutasjoner får æren for oppbygningen av biologiens orden og genetiske informasjon. I virkeligheten virker mutasjonene ensidig og irreversibelt nedbrytende på biologisk orden og genetisk informasjon.  At noen mennesker tenker slik, får bli deres sak. Problemet er når dette skal gjøres til samfunnets monopoliserte lære. Darwinismens framtredende holdning er at det ikke kan være legitimt å tenke eller lære annerledes i skole og kirke. Om folk som lærer annerledes får sparken, vitner det om at en ikke har mye toleranse mot annerledes-tenkende. Overfor kritikk kan darwinist-tilhengere opptre som de både sitter med fasit og kan sette sluttstrek samtidig. Innen riksdekkende media som NRK har darwinister i praksis hatt monopol så lenge at nykommere som bidrar med nytt budskap, anses som uvelkomne 'nybegynnere og fremmedelementer'.
At noen mennesker tenker slik, får bli deres sak. Problemet er når dette skal gjøres til samfunnets monopoliserte lære. Darwinismens framtredende holdning er at det ikke kan være legitimt å tenke eller lære annerledes i skole og kirke. Om folk som lærer annerledes får sparken, vitner det om at en ikke har mye toleranse mot annerledes-tenkende. Overfor kritikk kan darwinist-tilhengere opptre som de både sitter med fasit og kan sette sluttstrek samtidig. Innen riksdekkende media som NRK har darwinister i praksis hatt monopol så lenge at nykommere som bidrar med nytt budskap, anses som uvelkomne 'nybegynnere og fremmedelementer'.
 Media er kanskje den lydigste medløperen i så måte. Darwinistene har enkelte servilt underdanige tjenere i media. Kanskje den lydigste, er avisen som påberoper seg de sterkeste meningene: Dagbladet. Oss bekjent har det skjedd én gang at ei stakkars dame fikk spalteplass til et motinnlegg mot darwinismen. Det ble raskt fulgt opp av 4-5 darwinistiske motinlegg mot damen som ikke kom til med gjensvar. I slutten av en debatt med kun forsvar for darwinismen, sendte hovedansvarlig redaktør her inn motsvar. Han fikk til svar at 'debatt-festen var over'. Faktisk er det lettere å komme til med kritisk stoff mot den etablerte lære i Klassekampen. I det hele er det en mer åpen holdning overfor darwinismen i tidligere kommunist-land (eks. Tsjekkia) enn det er i Norge i dag.
Media er kanskje den lydigste medløperen i så måte. Darwinistene har enkelte servilt underdanige tjenere i media. Kanskje den lydigste, er avisen som påberoper seg de sterkeste meningene: Dagbladet. Oss bekjent har det skjedd én gang at ei stakkars dame fikk spalteplass til et motinnlegg mot darwinismen. Det ble raskt fulgt opp av 4-5 darwinistiske motinlegg mot damen som ikke kom til med gjensvar. I slutten av en debatt med kun forsvar for darwinismen, sendte hovedansvarlig redaktør her inn motsvar. Han fikk til svar at 'debatt-festen var over'. Faktisk er det lettere å komme til med kritisk stoff mot den etablerte lære i Klassekampen. I det hele er det en mer åpen holdning overfor darwinismen i tidligere kommunist-land (eks. Tsjekkia) enn det er i Norge i dag. Likevel ser forbausende mange nordmenn gjennom ensidigheten i media. De fleste later imidlertid til å tenke at: i) ensidigheten i media beror på enighet om sak. ii) Darwinistisk ensidighet i norske media er ikke tilsiktet, men følger av saklig enighet i akademisk verden. Media har jo nese for konflikter, og bør åpne opp for debatt på like premisser. Akademikerne lener seg mer på media enn omvendt. Det kan bli spennende når mediafolk oppdager at de ikke skylder darwinister noe, og at darwinister står i ensidig takknemlighetsgjeld til mediefolk -uten å yte gjengjeld. Om evolusjonskritikken skulle øke i media, skal det ikke så mye til før politikerne begynner gi darwinistene 95% i stedet for 100% av forskningsmidlene. Det er til sammenligning flere enn 5% som er helt mot darwinismen.
Likevel ser forbausende mange nordmenn gjennom ensidigheten i media. De fleste later imidlertid til å tenke at: i) ensidigheten i media beror på enighet om sak. ii) Darwinistisk ensidighet i norske media er ikke tilsiktet, men følger av saklig enighet i akademisk verden. Media har jo nese for konflikter, og bør åpne opp for debatt på like premisser. Akademikerne lener seg mer på media enn omvendt. Det kan bli spennende når mediafolk oppdager at de ikke skylder darwinister noe, og at darwinister står i ensidig takknemlighetsgjeld til mediefolk -uten å yte gjengjeld. Om evolusjonskritikken skulle øke i media, skal det ikke så mye til før politikerne begynner gi darwinistene 95% i stedet for 100% av forskningsmidlene. Det er til sammenligning flere enn 5% som er helt mot darwinismen. Praktisering av darwinismen i samfunnet
Praktisering av darwinismen i samfunnet
 samme svar som kommer tre ganger: 'Darwin med sin lære om mutasjoner og utvalg'. Menneskehetens tre eksistensielle spørsmål påkaller bare snusfornuftige skuldertrekk fra en darwinist. Svarene som en typisk ortodoks darwinist er i stand til å gi på disse tre er bare utrolig grunne og forutsigbare:
samme svar som kommer tre ganger: 'Darwin med sin lære om mutasjoner og utvalg'. Menneskehetens tre eksistensielle spørsmål påkaller bare snusfornuftige skuldertrekk fra en darwinist. Svarene som en typisk ortodoks darwinist er i stand til å gi på disse tre er bare utrolig grunne og forutsigbare:(Fra 'Evolutionens ikoner'+..; Jonathan Wells; Forlaget Origo)
 Men som vi har sett representerer evolusjonens ikoner i stedet en fordreid virkelighet av forskningsmaterialet. Tre av ikonene (evolusjons-stamtreet, fossile hester og menneskets opprinnelse) er legemliggjørelse av forestillinger som gir seg ut for å være nøytrale beskrivelser av naturen. To av ikonene (Haeckels fostre og bjørkemålerne) bygger på forfalskninger: Fosterbildene får virveldyrfostre til å se mer like ut enn de er i virkeligheten. Det er simpelten ikke sant at virveldyrfostre likner mest på hverandre på de tidsligste fosterstadier. Den firvingede bananfluen gir inntrykk av å være en mekanisme som kan fungere som evolusjonens råmateriale, men består av håpløst 'vingeskutt' insekt, en blindgate i kostbar forskningssammenheng.sensasjon om oppdagede falsknerier. Lærebokforfatter Bob Ritter brukte bevisst oppstilte bilder av bjørkemålere på trestammer. Han forsvarte sin handling med at han illustrerte et grunnleggende prinsipp. Så blir spørsmålet: 'er det i orden å illustrere et prinsipp en mener sant, med falsk dokumentasjon?' Noe av det samme gjelder for 'Bambiraptor', dinosauren som ble forsynt med fjær 'av teoretiske årsaker'. Da er det snakk om å presse observasjonene inn i en form. Slike saker har potensielt alvorlige konsekvenser for biologien.
Men som vi har sett representerer evolusjonens ikoner i stedet en fordreid virkelighet av forskningsmaterialet. Tre av ikonene (evolusjons-stamtreet, fossile hester og menneskets opprinnelse) er legemliggjørelse av forestillinger som gir seg ut for å være nøytrale beskrivelser av naturen. To av ikonene (Haeckels fostre og bjørkemålerne) bygger på forfalskninger: Fosterbildene får virveldyrfostre til å se mer like ut enn de er i virkeligheten. Det er simpelten ikke sant at virveldyrfostre likner mest på hverandre på de tidsligste fosterstadier. Den firvingede bananfluen gir inntrykk av å være en mekanisme som kan fungere som evolusjonens råmateriale, men består av håpløst 'vingeskutt' insekt, en blindgate i kostbar forskningssammenheng.sensasjon om oppdagede falsknerier. Lærebokforfatter Bob Ritter brukte bevisst oppstilte bilder av bjørkemålere på trestammer. Han forsvarte sin handling med at han illustrerte et grunnleggende prinsipp. Så blir spørsmålet: 'er det i orden å illustrere et prinsipp en mener sant, med falsk dokumentasjon?' Noe av det samme gjelder for 'Bambiraptor', dinosauren som ble forsynt med fjær 'av teoretiske årsaker'. Da er det snakk om å presse observasjonene inn i en form. Slike saker har potensielt alvorlige konsekvenser for biologien. Dogmatiske darwinister går ut med en snever måte å drive vitenskap på, og erklærer den for å være den eneste rette. Kritikerne får derpå benevnelsen 'uvitenskapelige', og deres artikler avvises av ledende tidskrifter hvor redaksjonen domineres av nettopp dogmatiske darwinister. Etter 'kollegialt gjennomsyn' blir kritikerne jaget ut av det vitenskapelige fellesskap. Derved oppnår en å minke i betydelig grad de forskningsresultater som går mot darwinismen. Evt. drukner en dem i lite tilgjengelige publikasjoner, hvor bare de mest iherdige kan finne dem igjen. Både i USA og Vest-Europa har en langt på vei lykkes i å lage en monopollignende kontroll over forskningsmidler, fakultetsfasiliteter og kollegiale publikasjoner. Om noen våger bryte ut av konsensusen, blir det et voldsomt oppstyr og vedkommende mister oftest jobben (eks. William Dembski).
Dogmatiske darwinister går ut med en snever måte å drive vitenskap på, og erklærer den for å være den eneste rette. Kritikerne får derpå benevnelsen 'uvitenskapelige', og deres artikler avvises av ledende tidskrifter hvor redaksjonen domineres av nettopp dogmatiske darwinister. Etter 'kollegialt gjennomsyn' blir kritikerne jaget ut av det vitenskapelige fellesskap. Derved oppnår en å minke i betydelig grad de forskningsresultater som går mot darwinismen. Evt. drukner en dem i lite tilgjengelige publikasjoner, hvor bare de mest iherdige kan finne dem igjen. Både i USA og Vest-Europa har en langt på vei lykkes i å lage en monopollignende kontroll over forskningsmidler, fakultetsfasiliteter og kollegiale publikasjoner. Om noen våger bryte ut av konsensusen, blir det et voldsomt oppstyr og vedkommende mister oftest jobben (eks. William Dembski). Holdningen en møter hos slike dogmatiske darwinister kan eksemplifiseres ved professor Jay Losey, valgt formann for Baylor universitets konsortium: "Hvis en avviser eller forkleiner evolusjonslæren, så stiller en spørsmålstegn ved hele den moderne vitenskaps grunnlag." I stedet for å forsvare akademisk frihet, prøver en knekke den. Ledere i NCSE (National Center for Science Education) står bak framstillinger av darwinistiske ikoner som faller nettopp i de feller vi har vist om deres ikoner her. Om noen prøver få alternative undervisningsmidler med inspirasjon fra Intelligent Design (ID), så nektes de dette. De støttes av ACLU, en organisasjon som skulle være opptatt av borgeres rettigheter, men synes mer interessert i å forsvare dogmatisk darwinismes privilegier. Også her i Norge går hundrevis av millioner skattepenger til darwinistisk basert forskning (Forskning og økonomi).
Holdningen en møter hos slike dogmatiske darwinister kan eksemplifiseres ved professor Jay Losey, valgt formann for Baylor universitets konsortium: "Hvis en avviser eller forkleiner evolusjonslæren, så stiller en spørsmålstegn ved hele den moderne vitenskaps grunnlag." I stedet for å forsvare akademisk frihet, prøver en knekke den. Ledere i NCSE (National Center for Science Education) står bak framstillinger av darwinistiske ikoner som faller nettopp i de feller vi har vist om deres ikoner her. Om noen prøver få alternative undervisningsmidler med inspirasjon fra Intelligent Design (ID), så nektes de dette. De støttes av ACLU, en organisasjon som skulle være opptatt av borgeres rettigheter, men synes mer interessert i å forsvare dogmatisk darwinismes privilegier. Også her i Norge går hundrevis av millioner skattepenger til darwinistisk basert forskning (Forskning og økonomi).
 Det foreligger likevel en fare ved en folkelig oppstand mot det darwinistiske etablissement ved at en risikerer kaste barnet ut med badevannet. Det er ikke forskningen som sådan det er noe feil ved. Offentlig finansiert forskning av høy kvalitet er av stor betydning for framtiden til velferdssamfunnet. Om fusk og fordreininger innen biologien skulle bidra til redusert tillit til forskning generelt, ville det være en tragedie. Det er grunnen til at sannhetssøkende, framfor dogmatiske, biologer formentlig vil overta føringen og renske opp i eget hus. På det viset kan de også unngå at en dogmatikk erstattes av en annen. Nettopp å spille på frykten for religiøs fundamentalisme, har vært meget effektiv for at de kan beholde sitt monopol. Ironisk nok 'beskytter' disse mennesker vitenskapen fra en type dogmatikk ved å påtvinge den deres egen. Den beste måten å renske opp på ville være å reetablere biologisk vitenskap på empirisk forskning, som er dens opprinnelige sanne grunnlag.
Det foreligger likevel en fare ved en folkelig oppstand mot det darwinistiske etablissement ved at en risikerer kaste barnet ut med badevannet. Det er ikke forskningen som sådan det er noe feil ved. Offentlig finansiert forskning av høy kvalitet er av stor betydning for framtiden til velferdssamfunnet. Om fusk og fordreininger innen biologien skulle bidra til redusert tillit til forskning generelt, ville det være en tragedie. Det er grunnen til at sannhetssøkende, framfor dogmatiske, biologer formentlig vil overta føringen og renske opp i eget hus. På det viset kan de også unngå at en dogmatikk erstattes av en annen. Nettopp å spille på frykten for religiøs fundamentalisme, har vært meget effektiv for at de kan beholde sitt monopol. Ironisk nok 'beskytter' disse mennesker vitenskapen fra en type dogmatikk ved å påtvinge den deres egen. Den beste måten å renske opp på ville være å reetablere biologisk vitenskap på empirisk forskning, som er dens opprinnelige sanne grunnlag. I 1973 uttalte neodarwinisten Theodosius Dobzhansky: "Intet innenfor biologien gir mening unntatt i lys av evolusjon." Dette har vært slagordet siden, for dogmatiske evolusjonister. For å rettferdiggjøre sine påstander bruker en typisk forskningsmateriale fra antibiotika -og penicillin-resistens, og mindre modifikasjoner innen artene. Som en konsekvens av dette hevdes at økonomisk avgjørende områder innenfor medisin og landbruk avhenger av deres innsats. For å behandle en infeksjon trengs ikke evolusjonsteori, men biokjemiske tester, for å avgjøre hvilke antibiotika som vil være best. Leger og landbruksfolk går gjerne pragmatisk til verks, og løser problemer etter som de oppstår. At en kan drive fram og forbedre avl på dyrs og planters egenskaper var kjent før Darwin. Vitenskap er ikke synonymt med darwinisme, som noen dogmatiske darwinister vil ha oss til å tro.
I 1973 uttalte neodarwinisten Theodosius Dobzhansky: "Intet innenfor biologien gir mening unntatt i lys av evolusjon." Dette har vært slagordet siden, for dogmatiske evolusjonister. For å rettferdiggjøre sine påstander bruker en typisk forskningsmateriale fra antibiotika -og penicillin-resistens, og mindre modifikasjoner innen artene. Som en konsekvens av dette hevdes at økonomisk avgjørende områder innenfor medisin og landbruk avhenger av deres innsats. For å behandle en infeksjon trengs ikke evolusjonsteori, men biokjemiske tester, for å avgjøre hvilke antibiotika som vil være best. Leger og landbruksfolk går gjerne pragmatisk til verks, og løser problemer etter som de oppstår. At en kan drive fram og forbedre avl på dyrs og planters egenskaper var kjent før Darwin. Vitenskap er ikke synonymt med darwinisme, som noen dogmatiske darwinister vil ha oss til å tro. påstanden om at "Intet innenfor biologien gir mening unntatt i lys av evolusjon," er altså påviselig falsk! Ja, faktisk kan den være en bedre biolog som avviser en slik påstand enn den som ukritisk godtar den. Naturvitenskapens største dyd er at den holder seg til forskningsmaterialet, får vi vite. Den som begynner med en forut-fattet idé , og ubevisst eller bevisst bøyer forskningsmaterialet i retning av det dyrker noe motsatt av forskning. Det er nettopp det Dobzhanskys maksime oppfordrer folk til å gjøre.
påstanden om at "Intet innenfor biologien gir mening unntatt i lys av evolusjon," er altså påviselig falsk! Ja, faktisk kan den være en bedre biolog som avviser en slik påstand enn den som ukritisk godtar den. Naturvitenskapens største dyd er at den holder seg til forskningsmaterialet, får vi vite. Den som begynner med en forut-fattet idé , og ubevisst eller bevisst bøyer forskningsmaterialet i retning av det dyrker noe motsatt av forskning. Det er nettopp det Dobzhanskys maksime oppfordrer folk til å gjøre.
(Fra 'Evolutionens ikoner'; Jonathan Wells; Forlaget Origo)
Faktum er at Miller-Urey forsøket antagelig ikke er en troverdig etterlikning av jordas tidlige atmosfære. Forsøket viser heller ikke hvordan livets kjemiske råmaterialer kan ha blitt til.Faktum er at hvis homologi defineres som likheter der skyldes felles avstamning, kan det ikke samtidig brukes som argument for samme felles avstamning. Hva enn årsaken kan være til at homologi opptrer mellom ulike dyreklasser, så har det i hvert fall ikke å gjøre med ensartede gener.Fakta om bananfluen Fakta om menneskets opprinnelse
Fakta om menneskets opprinnelseLes mer
 (fra Evolusjonens ikoner av Jonathan Wells; Origo forlag)
(fra Evolusjonens ikoner av Jonathan Wells; Origo forlag)
 evolusjonære stamtre. For 10 år siden håpet evolusjonister at det molekylære forskningsmaterialet ville kunne redde treet, men nyere oppdagelser har fått det håpet til å rakne. Selv om ikke biologibøkene sier det, er Darwins evolusjonstre revet opp med roten. 'Livets mektige tre' (opprinnelig tegning finnes ikke i høyere oppløsning).
evolusjonære stamtre. For 10 år siden håpet evolusjonister at det molekylære forskningsmaterialet ville kunne redde treet, men nyere oppdagelser har fått det håpet til å rakne. Selv om ikke biologibøkene sier det, er Darwins evolusjonstre revet opp med roten. 'Livets mektige tre' (opprinnelig tegning finnes ikke i høyere oppløsning). I følge Darwins teori har menneske og bananflue hatt samme stamfar.
I følge Darwins teori har menneske og bananflue hatt samme stamfar.
 I Afrika og Australia har geologer rapportert om sedimentbergarter som er over tre milliarder år gamle og inneholder fossiler fra éncellede organismer. En har funnet litt yngre sedimentbergarter, som inneholder fossile 'stromatolitter', lagdelte puteformede strukturer av fotosyntetiske bakteriesamfunn. Det er funnet spesielle flercellede organismer tidligere enn Kambrium, først funnet i Ediacara Hills i Syd-Australia, men senere flere steder. Noen paleontologer hevder at Ediacara-fossilene er forfedre til dyr som senere oppsto i Kambrium. Andre hevder at de er så ulike at de bør plasseres i sitt eget rike.
I Afrika og Australia har geologer rapportert om sedimentbergarter som er over tre milliarder år gamle og inneholder fossiler fra éncellede organismer. En har funnet litt yngre sedimentbergarter, som inneholder fossile 'stromatolitter', lagdelte puteformede strukturer av fotosyntetiske bakteriesamfunn. Det er funnet spesielle flercellede organismer tidligere enn Kambrium, først funnet i Ediacara Hills i Syd-Australia, men senere flere steder. Noen paleontologer hevder at Ediacara-fossilene er forfedre til dyr som senere oppsto i Kambrium. Andre hevder at de er så ulike at de bør plasseres i sitt eget rike.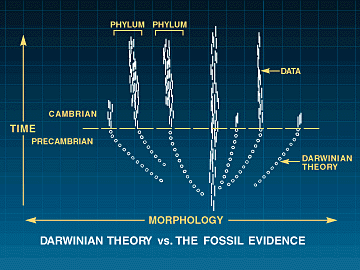 paleontolog -Jun-Yuan Chen. ( Paleontolog: en som forsker på bergarter, dyre- og plante-fossiler). Da han besøkte USA for å forelese om funnene, påpekte han at 'opp-ned' mønsteret fra Kambrium motsier darwinismen. Dette ble ignorert av forskerne der, og kommentert av gjesten slik: 'I Kina må en gjerne kritisere Darwin, men ikke regjeringen. I USA må man gjerne kritisere regjeringen, men ikke Darwin.'
paleontolog -Jun-Yuan Chen. ( Paleontolog: en som forsker på bergarter, dyre- og plante-fossiler). Da han besøkte USA for å forelese om funnene, påpekte han at 'opp-ned' mønsteret fra Kambrium motsier darwinismen. Dette ble ignorert av forskerne der, og kommentert av gjesten slik: 'I Kina må en gjerne kritisere Darwin, men ikke regjeringen. I USA må man gjerne kritisere regjeringen, men ikke Darwin.'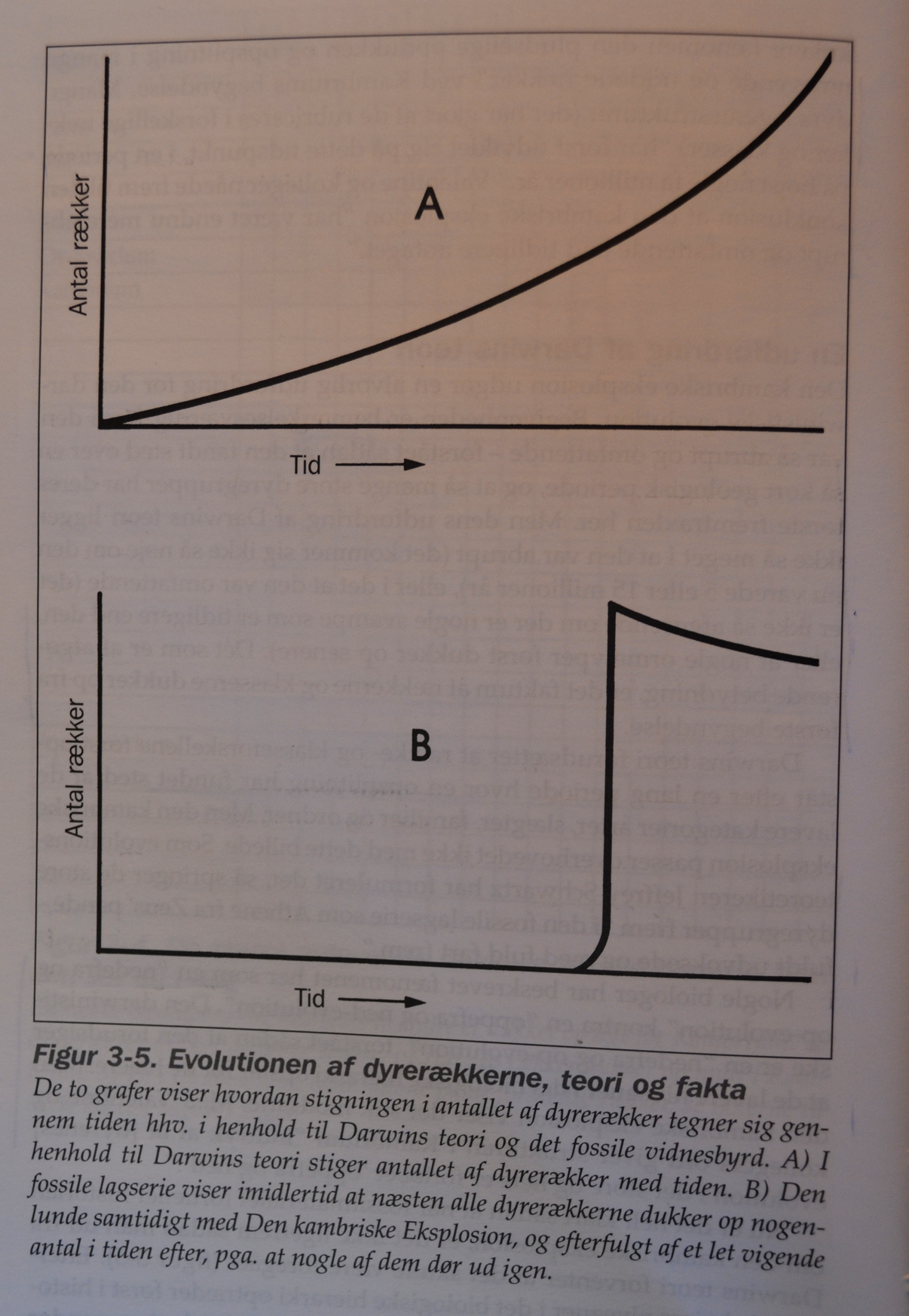 En utfordring til Darwins teori
En utfordring til Darwins teori Ad 1) En har i dag funnet tilstrekkelig gode sediment-bergarter fra sent PreKambrium og Kambrium til å overbevise paleontologene om at hvis det hadde fantes noen forfedre -og de hadde dannet fossiler, så ville de vært oppdaget nå. I følge James Valentine og Douglas Erwin er det slik at 'de serier av bergarter fra Kambrium vi har, er like så komplette og oppbygget under samme sedimentære forhold som lagserier som ligger er tettere på nåtiden. Allikevel er det slik at forfedre er "ukjente eller ubekreftede" for alle rekker eller klasser som dukket opp i den kambriske eksplosjon. "Den kambriske eksplosjon er ..for omfattende til at den kan bortforklares med defekter i den fossile lagserie." Flere ulike undersøkelser av den fossile lagserie fra Kambrium og opp til nå har bekreftet dette synet. I februar år 2000 når de britiske geologer M.J.Benton, M.A.Wills og R.Hitchin fram til flg.
Ad 1) En har i dag funnet tilstrekkelig gode sediment-bergarter fra sent PreKambrium og Kambrium til å overbevise paleontologene om at hvis det hadde fantes noen forfedre -og de hadde dannet fossiler, så ville de vært oppdaget nå. I følge James Valentine og Douglas Erwin er det slik at 'de serier av bergarter fra Kambrium vi har, er like så komplette og oppbygget under samme sedimentære forhold som lagserier som ligger er tettere på nåtiden. Allikevel er det slik at forfedre er "ukjente eller ubekreftede" for alle rekker eller klasser som dukket opp i den kambriske eksplosjon. "Den kambriske eksplosjon er ..for omfattende til at den kan bortforklares med defekter i den fossile lagserie." Flere ulike undersøkelser av den fossile lagserie fra Kambrium og opp til nå har bekreftet dette synet. I februar år 2000 når de britiske geologer M.J.Benton, M.A.Wills og R.Hitchin fram til flg.  konklusjon: "Den fossile lagseries tidligste deler er klart ufullstendige, men en kan saktens betrakte dem som pålitelige nok til å tegne de brede linjer i livets historie.
konklusjon: "Den fossile lagseries tidligste deler er klart ufullstendige, men en kan saktens betrakte dem som pålitelige nok til å tegne de brede linjer i livets historie.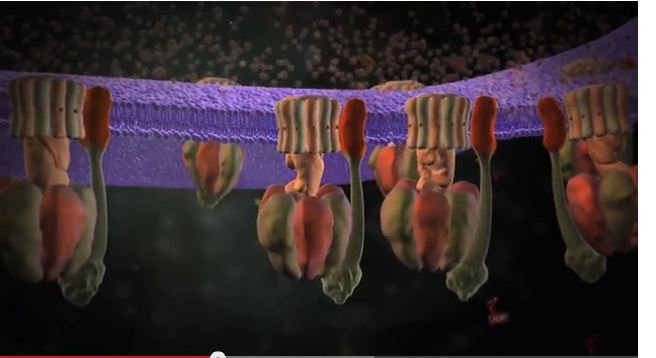 Ad 3) Den tredje måte evolusjonsbiologer har prøvd demontere den kambriske eksplosjon på, er ved å hevde at molekylært forskningsmateriale fra nålevende skapninger peker i retning av en felles stamfar til dyrerekkene, hundrevis av millioner år før Kambrium. For å forstå dette argumentet og hvorfor det ikke 'holder vann', er vi nødt til å se nærmere på 'molekylær fylogenese'
Ad 3) Den tredje måte evolusjonsbiologer har prøvd demontere den kambriske eksplosjon på, er ved å hevde at molekylært forskningsmateriale fra nålevende skapninger peker i retning av en felles stamfar til dyrerekkene, hundrevis av millioner år før Kambrium. For å forstå dette argumentet og hvorfor det ikke 'holder vann', er vi nødt til å se nærmere på 'molekylær fylogenese'  hvilke aminosyrer som inngår i en organismes mange ulike proteiner. Under replikasjonen blir rekkefølgen av baser kopiert til et nytt DNA-molekyl. Men molekylære tilfeldigheter eller mutasjoner kan en sjelden gang gjøre kopien ulik modermolekylet. Derfor kan organismer inneholde DNA-molekyler, og dermed også proteiner, som i noen grad avviker fra forfedrenes DNA og proteiner.
hvilke aminosyrer som inngår i en organismes mange ulike proteiner. Under replikasjonen blir rekkefølgen av baser kopiert til et nytt DNA-molekyl. Men molekylære tilfeldigheter eller mutasjoner kan en sjelden gang gjøre kopien ulik modermolekylet. Derfor kan organismer inneholde DNA-molekyler, og dermed også proteiner, som i noen grad avviker fra forfedrenes DNA og proteiner.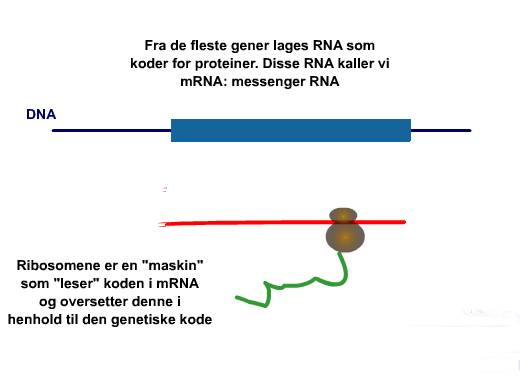 Da det ble raskere teknikker for analyse av DNA-sekvenser, ble det mer alminnelig å analysere genkodene enn proteinene, slik det tidligere var gjort. Utover DNA og proteiner, inneholder også alle organismer RNA, som er tett kjemisk beslektet med DNA. RNA bistår i å overføre informasjon fra DNA til proteinsekvensene. En del av denne prosessen beror på bittesmå organeller i cellen som kalles ribosomer. Se animasjon av oppbygningen her. Molekylær fylogenese og den kambriske eksplosjonnøyaktighet er stadig problematisk, i hvert fall for oppsplittingen av rekker". Grunnen for dette var at beregningene varierte med hundrevis av millioner år, avhengig av hvilke teknikker eller molekyler en bruker. Valentine og kolleger fastholder at "de molekylære data ikke kan rokke ved den [kambriske] eksplosjonen, som stadig framstår som en hovedbegivenhet i dyrenes evolusjon".
Da det ble raskere teknikker for analyse av DNA-sekvenser, ble det mer alminnelig å analysere genkodene enn proteinene, slik det tidligere var gjort. Utover DNA og proteiner, inneholder også alle organismer RNA, som er tett kjemisk beslektet med DNA. RNA bistår i å overføre informasjon fra DNA til proteinsekvensene. En del av denne prosessen beror på bittesmå organeller i cellen som kalles ribosomer. Se animasjon av oppbygningen her. Molekylær fylogenese og den kambriske eksplosjonnøyaktighet er stadig problematisk, i hvert fall for oppsplittingen av rekker". Grunnen for dette var at beregningene varierte med hundrevis av millioner år, avhengig av hvilke teknikker eller molekyler en bruker. Valentine og kolleger fastholder at "de molekylære data ikke kan rokke ved den [kambriske] eksplosjonen, som stadig framstår som en hovedbegivenhet i dyrenes evolusjon". Moderne versjoner av det darwinistiske Livets stamtre, kalles "fylogenetiske (stam)tre". På et typisk slikt tre, representerer "roten" stamformen til alle andre organismer i treet. De nederste greinene tegner de avstamningslinjer som forholdsvis tidlig har skilt seg ut, mens de høyere står for senere oppdelinger. Endene av grenene viser de egentlige arter. Oppdelingspunktet mellom to grener angir den hypotetiske stamformen for de to grenene. Mange slike trær tegnes slik at grenenes lengde svarer til forskjeller i sekvenser, som i sin tur skal indikere hvor lang tid det er gått siden de gikk hver til sitt (Fig3-7)Med andre ord: ulike molekyler fører til høyst ulike fylogenetiske stamtrær. Biologen Carl Woese en av pionerene bak konstruksjonen av rRNA-baserte stamtrær, kommenterte: "Inntil videre har ingen troverdige organisme-fylogeneser sett dagens lys, ut fra de mange individuelle protein-fylogeneser en har laget. Brudd på den fylogenetiske kontinuitet forekommer overalt i stamtreet. Det mangler sammenheng mellom roten og oppover. De større forgreininger henger ikke sammen. Gruppene innbyrdes gjør det heller ikke. Og dannelsen av de grunnleggende grupperinger henger heller ikke sammen.
Moderne versjoner av det darwinistiske Livets stamtre, kalles "fylogenetiske (stam)tre". På et typisk slikt tre, representerer "roten" stamformen til alle andre organismer i treet. De nederste greinene tegner de avstamningslinjer som forholdsvis tidlig har skilt seg ut, mens de høyere står for senere oppdelinger. Endene av grenene viser de egentlige arter. Oppdelingspunktet mellom to grener angir den hypotetiske stamformen for de to grenene. Mange slike trær tegnes slik at grenenes lengde svarer til forskjeller i sekvenser, som i sin tur skal indikere hvor lang tid det er gått siden de gikk hver til sitt (Fig3-7)Med andre ord: ulike molekyler fører til høyst ulike fylogenetiske stamtrær. Biologen Carl Woese en av pionerene bak konstruksjonen av rRNA-baserte stamtrær, kommenterte: "Inntil videre har ingen troverdige organisme-fylogeneser sett dagens lys, ut fra de mange individuelle protein-fylogeneser en har laget. Brudd på den fylogenetiske kontinuitet forekommer overalt i stamtreet. Det mangler sammenheng mellom roten og oppover. De større forgreininger henger ikke sammen. Gruppene innbyrdes gjør det heller ikke. Og dannelsen av de grunnleggende grupperinger henger heller ikke sammen.  En del molekylærbiologer mener problemet er metodologisk. I følge Forterre og Phillippe forandrer noen sekvenser seg for hurtig i evolusjonsmessig henseende til å bevare en "fylogenetisk melding" over lange tidsforløp. De hevder at ved å begrense seg til sekvenser som de mener har undergått en langsom evolusjon, kan de nå fram til et allmenngyldig stamtre. Problemet deres er at deres analyser peker i retning av en celle med kjerne som den altomfattende 'felles stamfar'. Tidligere er alltid darwinister gått ut fra at en celle uten kjerne er mer opprinnelig enn celler med kjerne. Ut fra darwinistisk synspunkt har 'treet slått rot på feil sted.'
En del molekylærbiologer mener problemet er metodologisk. I følge Forterre og Phillippe forandrer noen sekvenser seg for hurtig i evolusjonsmessig henseende til å bevare en "fylogenetisk melding" over lange tidsforløp. De hevder at ved å begrense seg til sekvenser som de mener har undergått en langsom evolusjon, kan de nå fram til et allmenngyldig stamtre. Problemet deres er at deres analyser peker i retning av en celle med kjerne som den altomfattende 'felles stamfar'. Tidligere er alltid darwinister gått ut fra at en celle uten kjerne er mer opprinnelig enn celler med kjerne. Ut fra darwinistisk synspunkt har 'treet slått rot på feil sted.'  En annen løsning på problemet er framsatt av W.Ford Doolittle, biolog ved Dalousie University. Han skriver i 1999: "Det kan godt være at det ennå ikke er lykkes .. å finne det riktige tre,..fordi livets historie i bunn og grunn ikke kan framstilles som et tre." I følge Doolittle skyldes de fylogenetiske uoverensstemmelser i overveiende grad "en gjensidig genoverføring." Doolittle lanserer den idé at genoverførsel mellom bakterier og urceller seg i mellom ligger til grunn for mange av uoverensstemmelsene i molekylære fylogeneser. Det innebærer at livets tidlige historie ikke ville likne et tre, men mer ett sammenflettet kratt (Fig3-8)evolusjonstreet stemmer ikke med de større trekk i det fossile og molekylære vitnesbyrd. Den kambriske eksplosjon viser at de høyeste kategorier er oppstått først, og det vender opp ned på Darwins evolusjonstre. (Se mer i "Darwins Doubt lenger ned i bloggen") Allikevel har "Livets Stamtre" stadig en dominerende plass blant evolusjonens ikoner, fordi darwinister har utropt det til å være et faktum!"det overveldende vitnesbyrd i Livets Stamtre er en realitet, og at ingen forsker så mye som ville drømme om å tvile på en altomfattende felles avstamning."
En annen løsning på problemet er framsatt av W.Ford Doolittle, biolog ved Dalousie University. Han skriver i 1999: "Det kan godt være at det ennå ikke er lykkes .. å finne det riktige tre,..fordi livets historie i bunn og grunn ikke kan framstilles som et tre." I følge Doolittle skyldes de fylogenetiske uoverensstemmelser i overveiende grad "en gjensidig genoverføring." Doolittle lanserer den idé at genoverførsel mellom bakterier og urceller seg i mellom ligger til grunn for mange av uoverensstemmelsene i molekylære fylogeneser. Det innebærer at livets tidlige historie ikke ville likne et tre, men mer ett sammenflettet kratt (Fig3-8)evolusjonstreet stemmer ikke med de større trekk i det fossile og molekylære vitnesbyrd. Den kambriske eksplosjon viser at de høyeste kategorier er oppstått først, og det vender opp ned på Darwins evolusjonstre. (Se mer i "Darwins Doubt lenger ned i bloggen") Allikevel har "Livets Stamtre" stadig en dominerende plass blant evolusjonens ikoner, fordi darwinister har utropt det til å være et faktum!"det overveldende vitnesbyrd i Livets Stamtre er en realitet, og at ingen forsker så mye som ville drømme om å tvile på en altomfattende felles avstamning." oppstått mer enn én gang og på ulike steder til ulike tider." Etter å ha fått tilgang til nyeste informasjon om molekylær fylogenese, skriver biologen Malcolm Gordon i 1998: "livet ser ut til å ha hatt atskillige opprinnelseskilder. Det universelle livets stamtre synes å ha mer enn bare en rot.. Den tradisjonelle versjon av teorien om felles avstamning, gjelder tilsynelatende ikke for riker ..[eller] rekker, og muligens heller ikke for mange av klassene innenfor rekkene.
oppstått mer enn én gang og på ulike steder til ulike tider." Etter å ha fått tilgang til nyeste informasjon om molekylær fylogenese, skriver biologen Malcolm Gordon i 1998: "livet ser ut til å ha hatt atskillige opprinnelseskilder. Det universelle livets stamtre synes å ha mer enn bare en rot.. Den tradisjonelle versjon av teorien om felles avstamning, gjelder tilsynelatende ikke for riker ..[eller] rekker, og muligens heller ikke for mange av klassene innenfor rekkene.Les mer
(Fra 'Evolutionens ikoner'; Jonathan Wells; Forlaget Origo)
 I 1840 årene kalte den britiske anatom Richard Owen den første form likheter for 'analogi' og den annen form for 'homologi'. Den gang ble skjelningen primært anvendt som hjelp til biologisk klassifisering. Analogi kan tyde på uavhengige tilpasninger til ytre forhold, mens homologi kan tyde på dypere strukturelt slektskap. Sistnevnte ble regnet for å være mest pålitelig når det gjaldt gruppering av organismer i familier, ordener, klasser og rekker.da benyttes til å argumentere for felles nedstamning. Det blir som å si at 'felles nedstamning skyldes felles nedstamning.' ii) Selv om det finnes enkelte felles gener med lik funksjonell virkning, har biologien mer blitt klar over at hver art har ulike epigenom, med kontekstavhengig informasjon for hver celletype. Dermed forblir mekanismen som skulle fremkalle homologi fortsatt ikke avklart!La oss betrakte eksempelet med forlemmers knokkelstrukturer (figur ovenfor) som Darwin mente var et vitnesbyrd om felles avstamning. En neodarwinist som vil avgjøre om virveldyrs forlemmer er homologe, må først avgjøre om de stammer fra en felles forfader eller ikke. Det må med andre ord være belegg for et felles opphav, før en kan kalle lemmer for homologe. Men hvis en så vender argumentasjonen rundt og argumenterer for at homologe lemmer peker på felles avstamning, blir det en grov sirkelslutning: Et felles opphav viser homologi, som i sin tur viser felles opphav. (Fig.4.2)
I 1840 årene kalte den britiske anatom Richard Owen den første form likheter for 'analogi' og den annen form for 'homologi'. Den gang ble skjelningen primært anvendt som hjelp til biologisk klassifisering. Analogi kan tyde på uavhengige tilpasninger til ytre forhold, mens homologi kan tyde på dypere strukturelt slektskap. Sistnevnte ble regnet for å være mest pålitelig når det gjaldt gruppering av organismer i familier, ordener, klasser og rekker.da benyttes til å argumentere for felles nedstamning. Det blir som å si at 'felles nedstamning skyldes felles nedstamning.' ii) Selv om det finnes enkelte felles gener med lik funksjonell virkning, har biologien mer blitt klar over at hver art har ulike epigenom, med kontekstavhengig informasjon for hver celletype. Dermed forblir mekanismen som skulle fremkalle homologi fortsatt ikke avklart!La oss betrakte eksempelet med forlemmers knokkelstrukturer (figur ovenfor) som Darwin mente var et vitnesbyrd om felles avstamning. En neodarwinist som vil avgjøre om virveldyrs forlemmer er homologe, må først avgjøre om de stammer fra en felles forfader eller ikke. Det må med andre ord være belegg for et felles opphav, før en kan kalle lemmer for homologe. Men hvis en så vender argumentasjonen rundt og argumenterer for at homologe lemmer peker på felles avstamning, blir det en grov sirkelslutning: Et felles opphav viser homologi, som i sin tur viser felles opphav. (Fig.4.2) forklaring på fenomenet. I USA kan det være vanskelig å finne fortalere for denne retningen, fordi det blant biologer er meget upopulært og risikabelt profesjonelt å sette spørsmålstegn ved Darwinismen.
forklaring på fenomenet. I USA kan det være vanskelig å finne fortalere for denne retningen, fordi det blant biologer er meget upopulært og risikabelt profesjonelt å sette spørsmålstegn ved Darwinismen. Hva viser DNA-sekvenser i retning av
Hva viser DNA-sekvenser i retning av er i morfologien." Så når de molekylære fylogeneser ikke stemmer overens, er eneste måteen kan skjelne mellom dem, at en vet noe om deres felles avstamning fra andre kilder -og herved er vi direkte tilbake i de sirkelslutninger som molekylære sammenlikninger skulle hjelpe oss komme ut av." (Fylogenese: utforsker slektskapsforholdene mellom arter eller grupper av arter. Ordet kommer av de greske røttene φύλον (fylon), «stamme» og γένεσις (genesis), «dannelse».)
er i morfologien." Så når de molekylære fylogeneser ikke stemmer overens, er eneste måteen kan skjelne mellom dem, at en vet noe om deres felles avstamning fra andre kilder -og herved er vi direkte tilbake i de sirkelslutninger som molekylære sammenlikninger skulle hjelpe oss komme ut av." (Fylogenese: utforsker slektskapsforholdene mellom arter eller grupper av arter. Ordet kommer av de greske røttene φύλον (fylon), «stamme» og γένεσις (genesis), «dannelse».) fossilenes evolusjonære slektskap som bygger på homologi som begrunnelse for felles avstamning " fanger oss hurtig inn i et nett av sirkelslutninger som det er umulig å unnslippe."
fossilenes evolusjonære slektskap som bygger på homologi som begrunnelse for felles avstamning " fanger oss hurtig inn i et nett av sirkelslutninger som det er umulig å unnslippe." Vidnesbyrd fra utviklingsretningerManglende sammenheng mellom homologi og utviklingsretninger, er gjeldende ikke bare i store trekk, men også i helt spesielle tilfeller ved virveldyrs lemmer. Det klassiske eksempel er salamanderne. I de fleste virveldyrs lemmer foregår utviklingen av fingre og tær bakfra og framover (fra hale mot hode), men hos salamanderne er det motsatt. Noen biologer har hevdet at salamandernes evolusjonshistorie må ha vært annerledes enn alle andre virveldyrs. Også når det gjelder utviklingen av virveldyrs skjelett-struktur, -som først formes som brusk og senere blir knokkelvev, er det uregelmessigheter:
Vidnesbyrd fra utviklingsretningerManglende sammenheng mellom homologi og utviklingsretninger, er gjeldende ikke bare i store trekk, men også i helt spesielle tilfeller ved virveldyrs lemmer. Det klassiske eksempel er salamanderne. I de fleste virveldyrs lemmer foregår utviklingen av fingre og tær bakfra og framover (fra hale mot hode), men hos salamanderne er det motsatt. Noen biologer har hevdet at salamandernes evolusjonshistorie må ha vært annerledes enn alle andre virveldyrs. Også når det gjelder utviklingen av virveldyrs skjelett-struktur, -som først formes som brusk og senere blir knokkelvev, er det uregelmessigheter: I følge neodarwinismen er det DNA-sekvenser eller gener som inneholder informasjon. Gener overfører informasjon fra generasjon til generasjon og styrer, i følge teorien, fosterets utvikling. Derfor er den teoretiske forklaring på homologe trekk at de er programmert av ensartede gener, nedarvet fra en felles forfader. Hvis en kan påvise at homologe strukturer i to forskjellige organismer er framkalt av tilsvarende gener, så ville vi ha forskningsmessig belegg for "informasjonskontinuiteten".Som eks. på dette anførte de Beer kun ét eksperiment som har med øyeutvikling hos bananfluen. Siden har en støtt på andre eksempler. Ett av dem er insekters leddannelse. Bananflue krever genet 'even-skipped' til utvikling av kroppsdeler, mens andre insekter utvikler ledd uten dette genet. Da alle insekt-ledd ansees være homologe, viser dette eksempel at homologe trekk ikke trenger være styrt av ensartede gener. Et annet eks. er 'sex-lethal', et gen som kreves til bestemmelse av kjønnet hos bananfluer, men ikke hos andre insekter..
I følge neodarwinismen er det DNA-sekvenser eller gener som inneholder informasjon. Gener overfører informasjon fra generasjon til generasjon og styrer, i følge teorien, fosterets utvikling. Derfor er den teoretiske forklaring på homologe trekk at de er programmert av ensartede gener, nedarvet fra en felles forfader. Hvis en kan påvise at homologe strukturer i to forskjellige organismer er framkalt av tilsvarende gener, så ville vi ha forskningsmessig belegg for "informasjonskontinuiteten".Som eks. på dette anførte de Beer kun ét eksperiment som har med øyeutvikling hos bananfluen. Siden har en støtt på andre eksempler. Ett av dem er insekters leddannelse. Bananflue krever genet 'even-skipped' til utvikling av kroppsdeler, mens andre insekter utvikler ledd uten dette genet. Da alle insekt-ledd ansees være homologe, viser dette eksempel at homologe trekk ikke trenger være styrt av ensartede gener. Et annet eks. er 'sex-lethal', et gen som kreves til bestemmelse av kjønnet hos bananfluer, men ikke hos andre insekter.. Mangel på overensstemmelse mellom gener og strukturer gjelder ikke bare for hele organismer, men også for lemmer. Et utviklingsgen som finnes i alle organismer, kalles 'Distal-less'. Det har fått dette navnet fordi en mutasjon i dette gen blokkerer bananfluers utvikling av lemmer (distal vedrører strukturer som ikke er en del av selve kroppen). Et gen med en ganske tilsvarende DNA-sekvens er funnet husmus. Gener som ligner 'Distal-less' er også funnet i sjøpiggsvin, børsteorm og ledd-orm lignende dyr (Onychophora).Konklusjonen står klar: Enten homologi skyldes nedarving med tilpasning eller ikke, forblir den spesifikke mekanismen bak det hele ukjent. Gavin de Beer skrev i 1971: "Hvilken mekanisme kan resultere i framstillingen av homologe organer, tross at de ikke styres av de samme gener? Jeg har stilt dette spørsmålet i 1938 og jeg har ennå ikke fått svar på det." Dette spørsmålet står fortsatt ubesvart.
Mangel på overensstemmelse mellom gener og strukturer gjelder ikke bare for hele organismer, men også for lemmer. Et utviklingsgen som finnes i alle organismer, kalles 'Distal-less'. Det har fått dette navnet fordi en mutasjon i dette gen blokkerer bananfluers utvikling av lemmer (distal vedrører strukturer som ikke er en del av selve kroppen). Et gen med en ganske tilsvarende DNA-sekvens er funnet husmus. Gener som ligner 'Distal-less' er også funnet i sjøpiggsvin, børsteorm og ledd-orm lignende dyr (Onychophora).Konklusjonen står klar: Enten homologi skyldes nedarving med tilpasning eller ikke, forblir den spesifikke mekanismen bak det hele ukjent. Gavin de Beer skrev i 1971: "Hvilken mekanisme kan resultere i framstillingen av homologe organer, tross at de ikke styres av de samme gener? Jeg har stilt dette spørsmålet i 1938 og jeg har ennå ikke fått svar på det." Dette spørsmålet står fortsatt ubesvart. homologi er felles avstamning, kan ikke homologi brukes som vitnesbyrd om felles avstamning'. Lærebøker i biologi prøver å underslå sirkelslutningen de gjør. Det er et problem for demokratiet. Samfunnet har bruk for folk som kan tenke selv, og ikke bare føyer seg etter hva eksperter og autoriteter forer dem med..
homologi er felles avstamning, kan ikke homologi brukes som vitnesbyrd om felles avstamning'. Lærebøker i biologi prøver å underslå sirkelslutningen de gjør. Det er et problem for demokratiet. Samfunnet har bruk for folk som kan tenke selv, og ikke bare føyer seg etter hva eksperter og autoriteter forer dem med..Les mer
(fra Evolusjonens IKONER, Vitenskap eller myte av Jonathan Wells)
 Darwin anså embryologien (læren om fosterutvikling) som det viktigste støttepunkt for sin teori. Når han skulle lage utkast til diverse dyreserier, har han i store trekk satt lit til embryologiske karakteristika. 'Det forekommer meg,' skrev han i Artenes Opprinnelse, 'at de viktigste kjennsgjerningene i embryologien (og ingen andre kommer på høyde med dem i betydning), kan forklares ut fra prinsippet om variasjoner i mange etterkommere fra en eller annen urstamfar'. Men da Darwin selv ikke var embryolog, var han helt avhengig av andres innsats på området. En av dem var den tyske professor Ernst Haeckel. Darwin skrev om ham at professor Haeckel bragte sin store viten og sine mange evner i anvendelse, med hensyn til noe han kaller fyliogenese (alle levende veseners avstamningslinjer).
Darwin anså embryologien (læren om fosterutvikling) som det viktigste støttepunkt for sin teori. Når han skulle lage utkast til diverse dyreserier, har han i store trekk satt lit til embryologiske karakteristika. 'Det forekommer meg,' skrev han i Artenes Opprinnelse, 'at de viktigste kjennsgjerningene i embryologien (og ingen andre kommer på høyde med dem i betydning), kan forklares ut fra prinsippet om variasjoner i mange etterkommere fra en eller annen urstamfar'. Men da Darwin selv ikke var embryolog, var han helt avhengig av andres innsats på området. En av dem var den tyske professor Ernst Haeckel. Darwin skrev om ham at professor Haeckel bragte sin store viten og sine mange evner i anvendelse, med hensyn til noe han kaller fyliogenese (alle levende veseners avstamningslinjer). Ja, Haeckel tegnet mange illustrasjoner. Den mest berømte er den med de tidlige virveldyrfostre. I følge Haeckels tegning er fostrene praktisk talt identiske på tidligste stadier, men blir så bemerkelsesverdig forskjellige etterhvert som de utvikler seg. 'Således er det sannsynlig, ut fra det vi vet om fostre hos pattedyr, fugler, fisk og krypdyr at disse dyr er modifiserte etterkommere fra en eller annen ur-stamfar', skrev Darwin. I sin bok 'The Descent of Man' utvidet Darwin sin deduksjon til også å gjelde mennesket.
Ja, Haeckel tegnet mange illustrasjoner. Den mest berømte er den med de tidlige virveldyrfostre. I følge Haeckels tegning er fostrene praktisk talt identiske på tidligste stadier, men blir så bemerkelsesverdig forskjellige etterhvert som de utvikler seg. 'Således er det sannsynlig, ut fra det vi vet om fostre hos pattedyr, fugler, fisk og krypdyr at disse dyr er modifiserte etterkommere fra en eller annen ur-stamfar', skrev Darwin. I sin bok 'The Descent of Man' utvidet Darwin sin deduksjon til også å gjelde mennesket.  poengterte imidlertid at 'de mer generelle trekk hos en stor del av dyrene kommer til syne tidligere i deres fostre enn de mer spesifikke trekk'.
poengterte imidlertid at 'de mer generelle trekk hos en stor del av dyrene kommer til syne tidligere i deres fostre enn de mer spesifikke trekk'.  Faktum er imidlertid at von Baer visste at fostre aldri ser ut som andre arters voksen-individer. Og han så ikke noe belegg for Darwins teori om at de ulike virveldyrklasser har samme stamfar. Von Baers to siste generaliseringer (lover) tok nettopp avstand fra at fosterutviklingen hvordan stamfaren til gruppa har sett ut. Selv om Darwin siterer von Baer som kilden til sitt argument fra embryologien, fordreier Darwin argumentet på avgjørende punkt for at det skal passe med sin teori. I det som vitenskapshistorikeren Fredrick Churchill kaller 'en de store ironiske ting ved det nittende århundres biologi,' ble von Baers synspunkt blandet sammen med og siden omdannet til en evolusjonær utgave av 'loven om parallellisme'. Naturalisten Fritz Müller bidro til dette, men det ble Müllers elev, Ernst Haeckel som gjorde forvirringen komplett.
Faktum er imidlertid at von Baer visste at fostre aldri ser ut som andre arters voksen-individer. Og han så ikke noe belegg for Darwins teori om at de ulike virveldyrklasser har samme stamfar. Von Baers to siste generaliseringer (lover) tok nettopp avstand fra at fosterutviklingen hvordan stamfaren til gruppa har sett ut. Selv om Darwin siterer von Baer som kilden til sitt argument fra embryologien, fordreier Darwin argumentet på avgjørende punkt for at det skal passe med sin teori. I det som vitenskapshistorikeren Fredrick Churchill kaller 'en de store ironiske ting ved det nittende århundres biologi,' ble von Baers synspunkt blandet sammen med og siden omdannet til en evolusjonær utgave av 'loven om parallellisme'. Naturalisten Fritz Müller bidro til dette, men det ble Müllers elev, Ernst Haeckel som gjorde forvirringen komplett. Til tross for at rekapitulasjonsteorien 'gikk av moten', forsøkte enkelte forskere (bl. a. Lillie) holde deler av den i live av 'tilpasnings grunner' i forhold til evolusjonsteorien. Men i 1922 ble den holdt for å være 'demonstrativt utroverdig' av den britiske embryolog Walter Garstang. Det var fordi 'de ontogenetiske trekk ikke bar på et eneste vitnesbyrd om de fullvoksne trekk som hører avstamningen til'. Garstang hadde ingen tillit til Haeckels hypotese om at nyutviklede trekk hekter seg som nye ledd på kjeden i fosterutviklins-prosessen: 'En hytte blir ikke et hus bare fordi en setter på en ekstra etasje ovenpå. Et hus representerer en høyere posisjon innen for boligutvikling, men det krever samtidig at hele bygningen bygges om.' (NB: dette må tolkes ut fra 1922 da hytter framdeles var hytter.)I diskusjoner om utvikling og evolusjon dukker to synspunkter til stadighet opp. Begge finnes i Darwins 'Artenes opprinnelse':
Til tross for at rekapitulasjonsteorien 'gikk av moten', forsøkte enkelte forskere (bl. a. Lillie) holde deler av den i live av 'tilpasnings grunner' i forhold til evolusjonsteorien. Men i 1922 ble den holdt for å være 'demonstrativt utroverdig' av den britiske embryolog Walter Garstang. Det var fordi 'de ontogenetiske trekk ikke bar på et eneste vitnesbyrd om de fullvoksne trekk som hører avstamningen til'. Garstang hadde ingen tillit til Haeckels hypotese om at nyutviklede trekk hekter seg som nye ledd på kjeden i fosterutviklins-prosessen: 'En hytte blir ikke et hus bare fordi en setter på en ekstra etasje ovenpå. Et hus representerer en høyere posisjon innen for boligutvikling, men det krever samtidig at hele bygningen bygges om.' (NB: dette må tolkes ut fra 1922 da hytter framdeles var hytter.)I diskusjoner om utvikling og evolusjon dukker to synspunkter til stadighet opp. Begge finnes i Darwins 'Artenes opprinnelse': sett. Likevel er de dukket opp som fugl Fønix av den empiriske avkreftelses-asken. Som vi skal se i det flg. illustreres begge med samme forfalskede tegninger.Men selv de fostre han har valgt ut, er forvansket for å kunne passe inn i hans teori. Den britiske embryolog Michael Richardson har i 1995 konstatert at : "Disse berømte bilder er upresise og gir et villedende bilde av fosterutviklingen. I 1997 la et ekspertpanel bestående av Richardson og et internasjonalt forskerteam Haeckels fostertegninger med fotografier av de riktige fostre fra alle syv klasser av virveldyr. Ut fra disse kan en tydelig se at Haeckels fostertegninger fortegner virkeligheten.
sett. Likevel er de dukket opp som fugl Fønix av den empiriske avkreftelses-asken. Som vi skal se i det flg. illustreres begge med samme forfalskede tegninger.Men selv de fostre han har valgt ut, er forvansket for å kunne passe inn i hans teori. Den britiske embryolog Michael Richardson har i 1995 konstatert at : "Disse berømte bilder er upresise og gir et villedende bilde av fosterutviklingen. I 1997 la et ekspertpanel bestående av Richardson og et internasjonalt forskerteam Haeckels fostertegninger med fotografier av de riktige fostre fra alle syv klasser av virveldyr. Ut fra disse kan en tydelig se at Haeckels fostertegninger fortegner virkeligheten.  Biologer har i over 100 år kjent til tidlige virveldyr-fostres ulikheter. Embryologen Adam Sedgwick understreket i 1894 at 'von Baers lov' om tidligere likheter ..ikke stemmer overens med det en kan slå fast om fosterutviklingen. Sedgwick sammenlignet fosteret av en liten hai med det til en høne og slo fast: 'Det finnes intet utviklingsstadium, hvorpå en så lett som ingenting kan se forskjell på dem med det blotte øye'.. 'Hvis von Baers lov skal gi mening, må den forutsette at dyr så nærbeslektet som høns og ender, ikke er til å skjelne fra hverandre på de tidligste stadier.. Ikke desto mindre kan jeg se forskjell på et høne og ande-embryo fra dag to" I følge Sedgwick kan "en art skjelnes fra alle de andre fra fosterutviklingens første stadier og hele veien opp."
Biologer har i over 100 år kjent til tidlige virveldyr-fostres ulikheter. Embryologen Adam Sedgwick understreket i 1894 at 'von Baers lov' om tidligere likheter ..ikke stemmer overens med det en kan slå fast om fosterutviklingen. Sedgwick sammenlignet fosteret av en liten hai med det til en høne og slo fast: 'Det finnes intet utviklingsstadium, hvorpå en så lett som ingenting kan se forskjell på dem med det blotte øye'.. 'Hvis von Baers lov skal gi mening, må den forutsette at dyr så nærbeslektet som høns og ender, ikke er til å skjelne fra hverandre på de tidligste stadier.. Ikke desto mindre kan jeg se forskjell på et høne og ande-embryo fra dag to" I følge Sedgwick kan "en art skjelnes fra alle de andre fra fosterutviklingens første stadier og hele veien opp."
 Et paradoks for darwinistisk utviklingsteori
Et paradoks for darwinistisk utviklingsteori Er et menneskefoster likt fisk?
Er et menneskefoster likt fisk? 'Gjeller' er ikke gjeller.
'Gjeller' er ikke gjeller. Når lærebokforfattere konfronteres med Haeckels falskneri for 100 år siden, kan en fortsatt oppnå å høre at en 'ikke visste om det'. F.eks. Douglas Futuyma (Evolutionary Biology 1998). Først etter å ha forhørt seg hos en utviklingsbiologi, hadde han fått kjennskap til Richardson og hans arbeid. Etter å ha lovet moderere dette i neste utgave, kommer han like fullt med påstanden om at fostrene er ganske like hverandre, bl.a. fordi de har gjeller! Noen biologer innrømmer at de har vært klar over dette i mer enn 20 år, men like fullt holdt stilt med det (eks: Stephen Jay Gould). Han har på sin side anklaget lærebokforfattere for å over-popularisere og nyhetsmediene for å skape sensasjon om stoffet. En kan lure på hvor tregt den selvkorrigerende effekten virker innenfor evolusjonsteorien. I flere tilfeller har det vært 'kreasjonister' som har kommet inn og påpekt feil (bl.a. Michael Behe).Slike fortielser/fordreininger har Gould selv kalt et 'akademisk sidestykke til mord'. Etter 100-års søvn burde være på tide å våkne opp!
Når lærebokforfattere konfronteres med Haeckels falskneri for 100 år siden, kan en fortsatt oppnå å høre at en 'ikke visste om det'. F.eks. Douglas Futuyma (Evolutionary Biology 1998). Først etter å ha forhørt seg hos en utviklingsbiologi, hadde han fått kjennskap til Richardson og hans arbeid. Etter å ha lovet moderere dette i neste utgave, kommer han like fullt med påstanden om at fostrene er ganske like hverandre, bl.a. fordi de har gjeller! Noen biologer innrømmer at de har vært klar over dette i mer enn 20 år, men like fullt holdt stilt med det (eks: Stephen Jay Gould). Han har på sin side anklaget lærebokforfattere for å over-popularisere og nyhetsmediene for å skape sensasjon om stoffet. En kan lure på hvor tregt den selvkorrigerende effekten virker innenfor evolusjonsteorien. I flere tilfeller har det vært 'kreasjonister' som har kommet inn og påpekt feil (bl.a. Michael Behe).Slike fortielser/fordreininger har Gould selv kalt et 'akademisk sidestykke til mord'. Etter 100-års søvn burde være på tide å våkne opp!
Les mer